 The
Nutcracker
The
Nutcracker 
![]() Kullstorlek
hos Nötkråka plus Summary.
Kullstorlek
hos Nötkråka plus Summary.
![]() Nötkråkans
vana att lägga upp vinterförråd.
Nötkråkans
vana att lägga upp vinterförråd.
![]() Ruvning
hos Nötkråkan
Ruvning
hos Nötkråkan
![]() Territory
in thick billed Nutcracker.
Territory
in thick billed Nutcracker.
![]() Food
Storage, Territory and Song in the Thick -
billed Nutcracker .
Food
Storage, Territory and Song in the Thick -
billed Nutcracker .
![]() Smalnäbbad
Nötkråka
Smalnäbbad
Nötkråka
 The hoarding habit of the Nutcracker Nucifraga c. caryocatactes.
The hoarding habit of the Nutcracker Nucifraga c. caryocatactes.
Summary below, meanwhile enjoy the pictures. By the way you will find text in English
with the pictures and tables. Just look for this sign
![]()
Nötkråkans äggläggning ett exempel på P O Swanbergs fantastiska forskningsinsatser vid sidan om kampen för Hornborgasjön. Längre ner på sidan följer en artikel som handlar om Nötkråkans fantastiska teknik att samla sitt vinterförråd. Ännu ett exempel på PO Swanbergs uppfinningsrikedom, envishet och noggrannhet.
Nötkråkans Nucifraga c. Caryocatactes vana att lägga upp vinterförråd ett annat fynd jag har gjort bland hans papper. The hoarding habit of the Nutcracker Nucifraga c. caryocatactes.
Kullstorlek hos Nötkråka Nucifraga caryocatactes i Skandinavien, relaterad till föregående års hasselnöttillgång
Per Olof Swanberg.


Alltsedan David Lack, med början 1947, i olika arbeten påvisade, hur äggkullens storlek hos många fåglar kan antagas vara beroende av tillgången till föda för blivande ungar, både hos arten som sådan och hos individer, har faktorer som kan påverka kullstorleken livligt diskuteras. Knappast hos någon annan art är relationen mellan aktuell födotillgång och kullens storlek tydligare än hos Nötkråkan.
I handböcker och specialarbeten han man i över hundra år läst, att kullens storlek hos Nötkråkan i Europa Nucifraga c. caryocatactes är 3 eller 4, någon gång 5 ägg. Otmar Reiser (1910), som sade sig basera sina erfarenheter på över 1000 ägg, som han "haft i sina händer", flertalet åren 1893-1898, uppgav, att antalet oftast är 3, mycket mer sällan än 4 och att han blott hade tre exemplar på 5- äggkullar, men att kullen mången gång utgjordes av 2 ägg. Makatsch (1976) anför:"3-4 ägg, tillfälligt 2, undantagsvis även 5 ägg... hittills har man ofta antagit, att kullstorleken vanligen skulle vara 3 ägg, detta skulle kunna sammanhänga med att man ofta insamlat kullar, som ej varit fullagda".
Egna erfarenheter av nötkråkstudier i granskogen på Billingen i Västergötland antydde efter några års arbete med artens häckningsbiologi, att kullens storlek kunde väntas vara relaterad till fågelparets tillgång till föda åt ungarna under dessas uppväxttid. Samtidigt hade det visat sig, att våra nötkråkor, som hade sina, uppenbarligen livsvariga, revir i djup barrskog, under hösten ägnar all sin tid och kraft åt att i mån av tillgång gömma vinterförråd i häckningsreviret, vilket på Billingen är 11-15 hektar stort. Minst ett nytt gömställe utnyttjades för varje transporterad börda. När ungarna kläckts, utgör nötkärnor ur föräldrarnas individuella förråd av hasselnötter deras huvudsakliga näring, se nedan ( även Swanberg 1951,1976).
Metodik
Eftersom nötkråkans häckningsbiologi och ekologi var föga känd, fördes anteckningar bl. a om äggantal. Anteckningar om hasselns fruktsättning har förts 1941 och 1943-1981 för att få underlag till bedömning av nötkråkornas möjlighet att insamla vinterförråd. Eftersom det är svårt att finna nötkråkornas bo - Reiser (1910) omtalade att man först letade förgäves i fem år - använde jag snart metoden att under vinter söndagar förse studieparen med nötter i deras respektive revir. Denna metod underlättade de allmänna studierna men medförde, att tyngdpunkten av egna anteckningar rörande kullstorlek kommit att representeras av nötkråkor som genom stödutfodring kunnat lägga nötter i förråd ända fram till äggläggningen. Det samma gäller observationer från Nils-Gerhard Karvik och Tore Lund, som länge sysslat med nötkråkestudier och med sina uppgifter bidragit till detta arbete. När man förser nötkråkor med nötter under vintern och våren, skyndar de sig nämligen att föra bort dessa, till dess att samtliga nötter gömts undan obehöriga blickar. För att få ett större antal uppgifter om kullstorleken hos nötkråkor som ej fått artificiellt tillskott av hasselnötter, har samlats övriga tillgängliga och tillräckligt distinkta uppgifter från Skandinaviens fastland.
Uppgifter från Mellan- och Sydeuropa har här ej medtagits, eftersom dessa ej alls kan relateras till tillgången till föda, lämplig för vinterförråd. I stora delar av Schweitz utgöres denna av frön av cembratall. Dennas frö är omgiven av ett motståndskraftigt skal, lämpar sig därför till förrådsuppläggning och kan f ö i evolutionshänseende betraktas som vår arts ursprungsföda. I t ex kantonen Graubünden förekommer, att nötkråkor har tillgång till och utnyttjar både cembrafrön och hasselnötter (Mattes 1978).
Äldre uppgifter från Ålands arkipelag (Rosenius 1949) och Bornholm (A. Benzon enl. Dresser 1871-81) har ej medtagits, i första hand därför att de ej är relaterade till nöttillgång och även därför att, enligt muntliga uppgifter av O. I. Karlsson, bosatt på ön Sottunga och erfaren av nötkråkor, hasselns fruktsättning i Ålands skärgård lär vara jämn och god från år till år. De jämnstora kullarna på öar vid Åland och i forna tider på Bornholm kan kanske förklaras med att dessa har ett maritimt präglat lokalklimat, som kan gynna hasselns fruktsättning. Viktigaste orsaken till utebliven fruktsättning uppgives nämligen vara frost vid den tid, då honblommorna utvecklas (Nils Sylvén, muntl.).
Här bör tilläggas, att de nakna fröna från våra inhemska barrträd av nötkråkan ej utnyttjas för vinterförråd. När grankottarna kränger i vårvintersol, kan man visserligen se nötkråkan plocka de frön, som fallit till snötäcket på marken och man kan också se dem plocka frön ur kottar i träden, men vid utfodringsförsök har mina nötkråkor synbarligen bara tagit frön för direkt konsumtion, ej visat tecken till att föra undan dem till förråd för framtida bruk.
Vinterförrådens betydelse för ungarna
De ovan omtalade vinterförråden har livsavgörande betydelse för ungarna i boet. Nötkråkan inleder häckningen under vinterförhållanden, och när ungarna har kläckts, utgör sönderdelade kärnor från de vinterförvarade nötterna deras huvudsakliga, under vissa år till en början troligen enda föda. Detta framhölls redan av Vogel (1873), och har bekräftats av direktobservationer under föräldrafåglarnas matningsprocedur på Billingen. N.G. Karvik fann vid undersökningen av två döda boungar, 2-3 veckor gamla, så sent som i mitten av maj 1947, att nötter i ungarnas magsäck var "alldeles dominerande mängd" (muntl. meddelande). Vidare fann D. Wikström "främst hasselnötter" i hakpåsen hos en hona, som var i färd med att mata en kull utflugna ungar den 20 maj 1927 (Palméns arkiv, Univ. Zool. Museum, Helsingfors)

Äggantalet styres av nötförrådens storlek
Den här anförda tabellen över huvudmaterialet (tabell) upptar dels 33 kullar hos nötkråkor som ej fått extra tillskott av nötter men lagt ägg under år, som följt på år med känd nöttillgång, dels 36 kullar, som lagts av stödutfodrade nötkråkpar. Därutöver redovisas, för fullständighetens skull, 8 kullar som följt på år med ej känd tillgång till hasselnötter. Slutsatsen av siffrorna är entydig: Om de under föregående höst samlade nötförråden blivit så begränsade, att tillgången till i förväg upplagrad föda för ungarna under dessas botid, d.v.s. 3-7 veckor efter äggläggningen, kan förutses bliva dålig eller snävt begränsad, lägger nötkråkan som regel endast 3 ägg. Om tillgången till föda för ungarna är rik (säkerställt tillräcklig) lägger nötkråkan normalt 4 ägg.
Tabell 1
Antal kullar No. of clutches |
|||||
| Hasselns fruktsättning föregående år Fruiting of the hazel in preceding autumn |
Efter regelbunden vintermatning med nötter. |
Okänd nöttillgång Hazel nut supply unknown |
|||
| Antal ägg Clutch size |
Dålig/svag Below average |
Genomsnittlig Average |
Rik Rich |
||
| 3 | 8 | 4 | 6 | 2 | 8 |
| 4 | 0 | 0 | 12 | 32 | 0 |
| 5 | 0 | 0 | 3 | 1 | 0 |
Tabell 1 Tjocknäbbad nötkråka Nucifraga c. caryocatactes. Fördelningen av kullar av olika storlek i relation till tillgången på hasselnötter i vinterförråd och i relation till utfodring med nötter under föregående vinter. Underlaget utgöres av material från specialstudium (69 kullar) och uppgifter från 8 andra bon, allt från Sveriges fastland 1936 - 1979.
![]() Thick - billed
Nutcracker. The size of clutches in relation to the available of hazel nuts stored in the
preceding autumn or during provisioning in the winter predecing breeding. Data on clutch
size come from 69 nests in a special study and from 8 other adequately checked nests, all
on Swedish mainland.
Thick - billed
Nutcracker. The size of clutches in relation to the available of hazel nuts stored in the
preceding autumn or during provisioning in the winter predecing breeding. Data on clutch
size come from 69 nests in a special study and from 8 other adequately checked nests, all
on Swedish mainland.
Skenbar avvikelse från regeln ?
Vid sammanställningen av material strävar man ofta efter att få så brett underlag som möjligt. För den skull sökte jag även efter uppgifter ur äggsamlingar. Resultatet blev överraskande och avvek markant från de ovan redovisade, som grundar sig på specialstudier. Det blev anledning att beakta David Davis (1955) ord beträffande problem vid insamlingen för forskningsändamål av data rörande kullstorlek: "data samlade från varierande källor är sällan tillräckligt användbara för specialundersökningar". För den skull behandlas dessa uppgifter för sig i tabellen 2 och 3. Orsaken till den överraskande skillnaden mellan å ena sidan resultatet av ett specialstudium, å andra sidan samlingar för annat ändamål är oklar. En tänkbar medverkande omständighet kan vara, att äggsamlingar är objekt i bytesverksamhet. Detta kan medföra förskjutningar, som ger en sned bild av verkligheten när det gäller fördelning av olika kullstorlek, bytesobjekt av olika attraktion. Det kan även tänkas, att vissa kullar "insamlats" innan de var fullagda.
Det förefaller ofruktbart att närmare spekulera över det förvånande materialet från äggsamlingar, när en specialundersökning visat, hur kullstorleken är reglerad efter klara linjer. Tabell tre visar, att man även efter sammanslagning av värdena från samtliga de 113 kullarna i tabellerna 1 och 2, som kunnat relateras till nöttillgång i vinterförråd, får resultat som visar kullstorlekens beroende av nöttillgången. För att ytterligare belysa vikten av Davis ord om vanskligheten av att basera undersökningsmaterial på blandade källor bör slutligen nämnas, att en sammanställning av sex kullar från fastlandet 1896-1879, nio från Bornholm 1863-1872 och tre från Ålands skärgård 1935-1937, alla i äggsamlingar, ger följande tal: 3-kullar = 39%, 4-kullar = 39% och 5-kullar = 22%! O. I. Karlsson, som sade sig ha, efter uppdrag av Paul Rosenius, letat upp sexton bon av nötkråka, förklarade vid mitt besök hos honom den 11 september 1949, att nästan alla hade innehållit 4 eller 5 ägg och att, som ovan redan anmärkts, hasselbuskarna på hans ö i skärgården alltid bar rikt med nötter, vilket kan ses som en naturlig förklaring till de stora kullarna. Vid mitt besök på Åland och kringliggande öar i sept. 1949 noterades, att hasselns fruktsättning där var rik - på Sveriges fastland hade jag det året registrerat "svag fruktsättning". Ytterligare ett exempel på vanskligheten i att använda blandade källor är O. I. Karlssons berättelse, att uppletandet av så många bon sammanhängde med att han trots ytterligt omsorgsfull packning av de till R sända äggen som regel fick beskedet, att de skadats i transporten och att därför nya kullar önskades.
Tabell 2
Antal kullar när hassels fruktsättning
under föregående höst var: |
||||
| Antal ägg i kullen Clutch size |
Dålig/svag Below average |
Genomsnittlig Average |
Rik Rich |
Okänd Unknown |
| 3 | 9 | 4 | 12 | 0 |
| 4 | 4 | 8 | 7 | 2 |
| 5 | 0 | 0 | 0 | 0 |
Tabell 2 Nötkråka i Sverige. Kullar under 1900-talet, noterade i olika äggsamlingar.
![]() The size of
Swedish clutches of the Nutcracker found in egg collections in relation to the hazel nut
crop of the autumn preceding their laying
The size of
Swedish clutches of the Nutcracker found in egg collections in relation to the hazel nut
crop of the autumn preceding their laying
5 ägg sällsynta undantag
Den sällsynta förekomsten av 5-äggkull har från Skandinaviens fastland i denna undersökning kunnat noteras endast 4 gånger i 123 kullar. Observera dock ovan nämnda siffror från äldre äggsamlingar ! De två gånger, då jag själv funnit så stor kull, inträffade detta hos ett och samma nötkråkpar, mitt par C. Detta i samband med det förhållandet att tre 5-kullar anträffats i den fåtaliga stammen på Bornholm under tre på varandra följande år, 1868-1870 (A. Benzon enl. Dresser 1871), kan antyda, att förmågan att under extremt gynnsamma förhållanden kunna lägga 5 ägg i en kull är bunden till vissa individer. Mitt bästa experimentpar A, som försågs med extra rikligt med nötter, mer än de övriga under åren 1941-1955, lade under dessa år 14 kullar med 4 ägg och en kull med 3, men aldrig någon med 5 trots den extremt rikliga nöttillgången och ytterligare tillskott av experimentföda som t.ex. möss talg. Tidigare, dvs. innan detta par hade börjat matas, hade det bara tre ägg i sitt år 1939 påträffade bo. Paret C hade på motsvarande sätt lagt 3 ägg 1941 och 1944 men lade kullar med 4 eller 5 ägg efter det rika nötåret 1941 och efter matning med nötter 1945-1947. Dessa exempel kan, som ovan nämnts, tas som indikation på att förmågan att lägg 5 ägg är genetiskt betingad. Haukioja (1970) kom i sin undersökning rörande sävsparven Emberiza Schoeniclus också till slutsatsen att övre gränsen för kullstorlek är genetiskt styrd. Yom-Tov (1974) drog av sina experiment slutsatsen, att kullstorleken hos kråkan Corvus corone är nedärvd.
Existerar 2- och 7 äggskullar ?
Såsom inledningsvis nämndes, skulle enligt äldre uppgifter kullar med endast två ägg kunna förekomma (Reiser 1910). Emellertid saknas sådana uppgifter från Skandinavien bland 144 redovisade kullar, trots vissa år med extrem brist på nötter. Man observerar, att sådana uppgifter härstammar från ett inledningsskede under intensiv äggsamlarverksamhet. I en serie av 42 kullar, som Reiser i detalj redovisat, har medtagits 2-äggkullar. Om den ena av dessa skrev Reiser själv: "sannolikt ofullständig kull". Det är säkert risk att ofullständiga kullar insamlas när man, som Reiser gjorde kring Sarajevo våren 1895, insamlar 18 kullar med hjälp av åtta medhjälpare från orten ! Någon kan ha haft större intresse av att medföra färska ägg till uppdragsgivaren än att låta dem ligga för förnyad kontroll av bo, till vilket man dessutom kanske ej skulle kunna återvända. Seriösa uppgifter om 2-äggkullar från forskare, som bemödat sig om att själva kontrollera, om kullen varit fullagd, synes saknas. Motsvarande gäller för gamla uppgifter om upp till 7 ägg hos nötkråkan. G Vogel avvisade sådana uppgifter redan 1873 och detta uppenbarligen med all rätt, eftersom större kull än 5 sedan ej iakttagits under mer än hundra år.
Nötförråden livsviktiga för avkomman
Nötkråkans tydliga reglering av kullstorleken är en god bekräftelse på David Lacks (1948,1954,1968) teori att kullstorleken hos fåglar brukar vara anpassad till ett med hänsyn till avkommans utveckling optimalt antal ungar. Den är ytterligare ett bekräftande exempel på motsvarande fynd och slutsatser i samma riktning, som beträffande kråka Corvus corone gjorts av Tenoveuo (1963), Loman (1977), Wittenberg(1968), men dock ej kunnat experimentellt bekräftas av Yom-Tov (1974). Lack grundade sin teori på olika iakttagelser av högre dödlighet hos ungar i större kullar under de första två månaderna. Hos Nötkråkan börjar den kritiska tiden för ungarna genast under de första veckorna i boet, då allt hänger på om föräldrarnas under hösten upplagda matförråd skall räcka. Ett dramatiskt exempel på detta gavs i april 1944. Föregående höst, 1943 hade hasselns fruktsättning varit extremt dålig. Jag hade i mina nötkråkors hasselmarker noterat "ungefär 1 nöt på 20 hasselbuskar". I nordvästra Skåne räknades under hösten 1943 samtliga nötter 32 st., på ett bestånd av 61 odlade buskar. Vid jämförelse med skörden på samma buskar under det nötrika året 1945 framgick att skörden 1943 förhöll sig till skörden 1945 som 1:450! En tydlig illustration av orsaken till den stora utvandringen av yngre nötkråkor hösten 1943. Men de gamla fåglar, som redan hade väletablerade revir på Billingen, stannade kvar i dessa efter att ha samlat vad de kunde av nötter. Under efterföljande vinter hade jag ännu inte lyckats förse mitt nötkråkepar B med nötter, och jag noterade att jag under den period, då marken var snötäckt, funnit "långt färre nötskal" hos detta par än i den andra tre reviren, som jag höll på med. Paret B började dock bygga i sin snöhöljda skog den 30 mars och hade sin 3-äggkull klar den 13 april 1944. Efter jämt 18 dygns ruvning kläcktes alla tre äggen inom 6 timmar, vilket visar, att de fått fullt normal embryonalutveckling.
Från en observationskoja kunde jag under tre dagslånga bevakningar iaktta, att föräldrarna matade ungarna mycket mindre än normalt. I början av maj var det snö i skogen och ingen annan mat att hämta än vad återstående nötförråd kunde ge. Föräldrarna lämnade ungarna oskyddade onormalt mycket, vilket tolkades som orsakat av förtvivlade försök att finna mer föda, ibland medförde de ingen mat alls vid avlösningsbesök i boet, vilket också är exceptionellt.
Nötkråkeungar ligger normalt tysta i boet, men dessa lät höra hungerskrik, som när ungar var 16 dagar gamla i lugnt väder kunde höras 70 meter bort i skogen; när ungarna var 21 dagar gamla, kunde hungerskriken höras på 160 meters håll. Sådana skrik från nötkråkungar i bo har jag varken förr eller senare upplevt. Jag försökte stödja föräldrarna genom att vid besök den knappa fritiden lägga granfrö på bokanten, men det var för sent. När ungarna var 16 dagar gamla, såg de små och magra ut, vid 21 dagar noterades, att de såg ut som "små dvärgar", en av dem var död, och en av de följande återstående dagarna hade de återstående två svultit ihjäl som en följd av att det varit omöjligt för föräldrarna att samla ihop tillräckligt för råd.
Trots en hel vinters katastrofsituation hade nötkråkan ej lagt mindre än 3 ägg. Det stöder åsikten att man ej bör åberopa obekräftade uppgifter om 2-äggskullar.
Vid studiet av frekvensen av respektive kullstorlek i relation till näringsförrådet är det en omständighet av särskilt intresse, nämligen att det är det individuella, under föregående år upplagda näringsförrådet, i sin tur beroende av föregående års fruktsättning hos värdväxten hasseln, som i Skandinavien avgör kullstorleken.

Nötkråkan matas med nötter
Ransoneras förrådens utnyttjande för eget bruk ?
Ett ytterst intressant förhållande är, att nötkråkan, som är beroende av vinterförråden även för egen förmåga att övervintra, efter svåra bristår har nötter kvar, när äggen kläckts vid mitten eller slutet av april. Så var det åtminstone i en del fall även efter den extrema nötbristen 1943. För min del kan jag inte avvisa tanken att föräldrafåglarna faktiskt under vintern håller igen på utnyttjandet av otillräckliga vinterförråd, eftersom de vanligen, trots allt, verkligen synes få ut sina ungar även under den svåra situationen efter bristår. Under år med svår nötbrist kan de bli nödsakade att sluta samla nötter redan i första veckan av september, dvs. efter 2-3 veckors besvärligt och ineffektivt letande. Andra år kan en flitig skytteltrafik med nötbördor mellan hasselängen och reviret i barrskogen pågå dagen lång ända in i december (Swanberg 1951,1976)
Hypotesen att en viss reglering av utnyttjandet av vinterförråden kan ske, utgår från följande två resonemang: (A) Om ransonering av de individuellt upplagda förråden inte skulle ske, borde nötkråkan under vinterperioden med ungefär 4 månaders snötäcke förväntas ta i anspråk en viss "standardkvantitet" per dag, en kvantitet som huvudsakligen skulle variera med temperaturen i reviret, eftersom nötkråkan stannar i detta under vintern, synbarligen ej utnyttjar granfrö förrän kottarna kränger i vårsolen och har starkt begränsade andra näringsmöjligheter, t.ex. näbbmöss som visar sig ovan snön. Utan ransonering skulle alltså vid svår nötbrist förråden vara tömda redan före äggläggningen. Även om alla honor lade 3-äggkullar, skulle då inga nötter alls finnas för ungarnas uppfödning och svältdöd följa. Efter medelmåttiga nötår skulle förråden kanske räcka fram till äggläggningen men ta slut strax därefter. Med därigenom oförändrad fett- och proteinreserv (jmf t.ex. Krapu 1981) kunde honan då förväntas lägga 4-äggkull, trots att föda till ungarna senare skulle saknas. Den reglering av äggantalet som sker är svårförklarlig, om man inte antar en ransonering av förråden. (B) Nötkråkor har ett väl fungerande abstrakt minne, som medför, att de ännu vid slutet av vinterperioden minns vilka nötgömmor, de utnyttjat, och därför ej riskerar att gräva efter förråd, som redan är tömda (Swanberg 1951,1976). Att de på detta sätt i cirka sju månader besparas ett förvirrat letande bland många hundra, kanske ett tusental gömställen, bör underlätta för fågeln att på något sätt registrera i vilken grad förråden utnyttjas. Ja, hennes minne är utomordentligt, att om hon efter ett mycket nötrikt år ej haft användning för nötterna ur alla gömställen, minns hon återstående sådana ännu efter ca 17 månader (Swanberg 1976). Efter det dåliga nötåret 1951 sågs sålunda en nötkråka den 27 och 31 mars och 6 april 1952 gräva upp ännu ej grodda men svartskaliga nötter, som fågeln hade kvar från ett överskott av hasselnötter från den rikare hösten 1950.
Den varelse, vars sinnesförmögenheter kan vara så subtilt inrättade, bör genom "förvärvat inbyggd" uppfattning om sina gömställens omfattning kunna reglera såväl eget utnyttjande av dem under vintern som äggantalet med hänsyn till storleken av återstående förråd, likaväl som olika andra fågelarter kan i viss mån anpassa kullstorleken efter rådande näringsförhållanden, som deras organism på något sätt kunnat registrera (jmf t.ex. gnagarberoende rovfåglar).
"Numerostatens" funktion
Erfarenheten visar, att om man väl känner nöttillgången i nötkråkornas insamlingsområde under augusti september, kan man med hög grad av sannolikhet förutsäga antalet ägg hos dem under följande häckningssäsong.
Man kan fundera vad som är normalt för den tjocknäbbade nötkråkan så, att fågeln i fysiologiskt avseende har ett kullstorleksmaximum vid 4 ägg, ett minimum vid 3 ägg. För att utnyttja ett av John Steen & Howard Paker 1981 lanserat begrepp: vid begränsade nötförråd ställer nötkråkan in ägg numerostaten på 3 ägg, vid rika nötår, som räcker för den kommande försörjningen av 4 ungar, är numerostaten inställd på 4 ägg. Som ovan nämnts är 5 ägg ett sällsynt individuellt undantag, troligen genetiskt betingat.
Mot en sådan bakgrund synes det ha en naturlig förklaring att von Brömssen & Jansson (1980) fann, att talltitan och tofsmesen ej ökade sin kullstorlek vid artificiell tillförsel av överskott av föda. Experimentobjekten kan antagas ha haft den fysiologiska kullkapaciteten som - enligt Lacks hypotes - naturen skänkt dem. Jämförelseparen har även utan extra tillförsel av föda kunnat utnyttja denna normala fysiologiska maximumkapacitet, vilket antydes av att deras äggantal väl stämmer med vad von Haartman (1969) anger som normalt för arterna. Experimentfåglarna bör, liksom nötkråkan, ej ha kunnat överskrida det fysiologiska maximumantalet, även om födotillskottet varit långt över det normala.
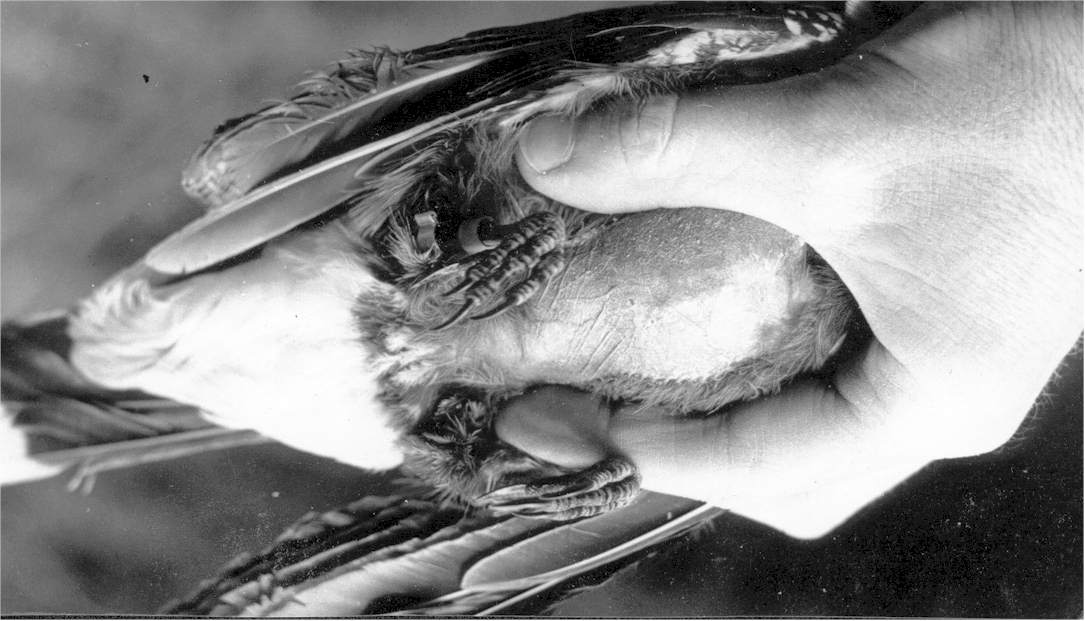
Nötkråkan finner sitt gömställe

Nötkråkan B har kommit till boet med mat förvarad i fågelns speciella, under tungan belägna hakpåse. Åtminstone på orginalbilden kan man genom näbbspringan skönja den gröt av fragment av nötkärnor, som stötts upp och med vilken nötkråkan matar sina 7 dagar gamla ungar den 7 maj 1944. Genom extrem nötbrist 1943 hade det under hösten anlagda förrådet av nötter blivit otillräckligt. Det räckte inte ens till försörjningen av tre ungar. När bilden togs, var ungarna redan hämmade i sin utveckling. De svalt ihjäl vid tre veckors ålder.
![]() The
Nutcracker feeding 7 days old young after a very poor hazel year of 1943. When
the picture
was taken on the 7th of May 1944, the growth of the young was already retarded. Due to
insufficient amounts of food in the parents winter stores the nestlings later starved to
death, three weeks old.
The
Nutcracker feeding 7 days old young after a very poor hazel year of 1943. When
the picture
was taken on the 7th of May 1944, the growth of the young was already retarded. Due to
insufficient amounts of food in the parents winter stores the nestlings later starved to
death, three weeks old.
Ransoneras förrådens utnyttjande för eget bruk ?
Ett ytterst intressant förhållande är, att nötkråkan, som är beroende av vinterförråden även för egen förmåga att övervintra, efter svåra bristår har nötter kvar, när äggen kläckts vid mitten eller slutet av april. Så var det åtminstone i en del fall även efter den extrema nötbristen 1943. För min del kan jag inte avvisa tanken att föräldrafåglarna faktiskt under vintern håller igen på utnyttjandet av otillräckliga vinterförråd, eftersom de vanligen, trots allt, verkligen synes få ut sina ungar även under den svåra situationen efter bristår. Under år med svår nötbrist kan de bli nödsakade att sluta samla nötter redan i första veckan av september, dvs. efter 2-3 veckors besvärligt och ineffektivt letande. Andra år kan en flitig skytteltrafik med nötbördor mellan hasselängen och reviret i barrskogen pågå dagen lång ända in i december. (Swanberg 1951,1976)
Hypotesen att en viss reglering av utnyttjandet av vinterförråden kan ske, utgår från följande två resonemang: (A) Om ransonering av de individuellt upplagda förråden inte skulle ske, borde nötkråkan under vinterperioden med ungefär 4 månaders snötäcke förväntas ta i anspråk en viss "standardkvantitet" per dag, en kvantitet som huvudsakligen skulle variera med temperaturen i reviret, eftersom nötkråkan stannar i detta under vintern, synbarligen ej utnyttjar granfrö förrän kottarna kränger i vårsolen och har starkt begränsade andra näringsmöjligheter, t.ex. näbbmöss som visar sig ovan snön. Utan ransonering skulle alltså vid svår nötbrist förråden vara tömda redan före äggläggningen. Även om alla honor lade 3-äggkullar, skulle då inga nötter alls finnas för ungarnas uppfödning och svältdöd följa. Efter medelmåttiga nötår skulle förråden kanske räcka fram till äggläggningen men ta slut strax därefter. Med därigenom oförändrad fett- och proteinreserv (jmf t.ex. Krapu 1981) kunde honan då förväntas lägga 4-äggkull, trots att föda till ungarna senare skulle saknas. Den reglering av äggantalet som sker är svårförklarlig, om man inte antar en ransonering av förråden. (B) Nötkråkor har ett väl fungerande abstrakt minne, som medför, att de ännu vid slutet av vinterperioden minns vilka nötgömmor, de utnyttjat, och därför ej riskerar att gräva efter förråd, som redan är tömda (Swanberg 1951,1976). Att de på detta sätt i cirka sju månader besparas ett förvirrat letande bland många hundra, kanske ett tusental gömställen, bör underlätta för fågeln att på något sätt registrera i vilken grad förråden utnyttjas. Ja, hennes minne är utomordentligt, att om hon efter ett mycket nötrikt år ej haft användning för nötterna ur alla gömställen, minns hon återstående sådana ännu efter ca 17 månader (Swanberg 1976). Efter det dåliga nötåret 1951 sågs sålunda en nötkråka den 27 och 31 mars och 6 april 1952 gräva upp ännu ej grodda men svartskaliga nötter, som fågeln hade kvar från ett överskott av hasselnötter från den rikare hösten 1950.
Den varelse, vars sinnesförmögenheter kan vara så subtilt inrättade, bör genom "förvärvat inbyggd" uppfattning om sina gömställens omfattning kunna reglera såväl eget utnyttjande av dem under vintern som äggantalet med hänsyn till storleken av återstående förråd, likaväl som olika andra fågelarter kan i viss mån anpassa kullstorleken efter rådande näringsförhållanden, som deras organism på något sätt kunnat registrera (jmf t.ex. gnagarberoende rovfåglar).

Numerostatens" funktion
Erfarenheten visar, att om man väl känner nöttillgången i nötkråkornas insamlingsområde under augusti september, kan man med hög grad av sannolikhet förutsäga antalet ägg hos dem under följande häckningssäsong.
Man kan fundera vad som är normalt för den tjocknäbbade nötkråkan så, att fågeln i fysiologiskt avseende har ett kullstorleksmaximum vid 4 ägg, ett minimum vid 3 ägg. För att utnyttja ett av John Steen & Howard Paker 1981 lanserat begrepp: vid begränsade nötförråd ställer nötkråkan in ägg numerostaten på 3 ägg, vid rika nötår, som räcker för den kommande försörjningen av 4 ungar, är numerostaten inställd på 4 ägg. Som ovan nämnts är 5 ägg ett sällsynt individuellt undantag, troligen genetiskt betingat.
Mot en sådan bakgrund synes det ha en naturlig förklaring att von Brömssen & Jansson (1980) fann, att talltitan och tofsmesen ej ökade sin kullstorlek vid artificiell tillförsel av överskott av föda. Experimentobjekten kan antagas ha haft den fysiologiska kullkapaciteten som - enligt Lacks hypotes - naturen skänkt dem. Jämförelseparen har även utan extra tillförsel av föda kunnat utnyttja denna normala fysiologiska maximumkapacitet, vilket antydes av att deras äggantal väl stämmer med vad von Haartman (1969) anger som normalt för arterna. Experimentfåglarna bör, liksom nötkråkan, ej ha kunnat överskrida det fysiologiska maximumantalet, även om födotillskottet varit långt över det normala.
Summary
![]() Please observe You will also find text by the pictures and tables
Please observe You will also find text by the pictures and tables
Clutch size in the Thick-billed Nutcracker, Nucifraga c. caryocatactes in Scandinavia in relation to the supply of hazel nuts in the individual winter stores.
This study was carried out mainly in the spruce forest on the Billingen Hills, 130 km NE of Gothenburg , southern Sweden. Adult Tick-billed Nutcrackers are resident and territorial. From late august until supplies are exhausted they spend most of the day caching hazel nuts from on their territory. Hazelnuts from the parent birds´ individual winter stores are of vital importance to the nestling and to the fledglings during the first weeks after leaving the nest. Hatching normally takes place in the third week of April, when the forest is often covered with snow.
The fruiting of the hazel varies considerably. In one district in South Sweden the rich crop of 1945 was 450 times that of the extremely poor year of 1943. (In 1944, the young in one Nutcracker nest starved to death, see fig above. The fruiting of the hazel, i e the availability of nuts , was recorded in 1941, and from 1943 until 1981. Some Nutcracker territories were supplied with extra nuts in winter. The "experimental" birds quickly hoarded the nuts presented. As shown in table 1 clutch size in the Thick-billed Nutcracker is clearly related to the availability of food (hazel nuts) stored. If the amount of hazel nuts hoarded in the preceding autumn is small, the Nutcracker as a rule will lay only three eggs. If the amount of food stored is sufficiently large to last over April - May, even for a lager brood, the Nutcracker will normally lay 4 eggs.
In this study only 5 out of 123 nests contain 5-egg clutches. The pair A, that was checked over 16 years, laid a clutch of 3 in 1939 (I g before I began to feed it). In later years, when the pair was provided nuts in winter, the female only once laid 3 eggs against 4 eggs in fourteen years. In spite of unusually rich provisioning that this pair experienced, the female never produced a clutch of 5 eggs. The pair C laid 3 eggs in 1941 and 1944; in 1942, after the rich hazel grow of 1941, and after the provision of food in 1945 to 1947, however, the female laid two 4-egg clutches and two 5-egg clutches. It is thought that the ability to lay 5 eggs is rare, individual, genetically dependent exception. It is suggested that the Thick-Billed Nutcracker has physiological minimum of 3 eggs, and a normal physiological maximum of 4 eggs. Old records of 7-egg clutches are more in doubt; the few 2-egg clutches of older reports were probably not completed. No 2-egg clutches were found among 144 Scandinavian clutches.
As shown above, the clutch of the Nutcracker in Scandinavia is clearly regulated by the fruiting of the hazel in the autumn preceding laying. This in turn determines the size of individual hazel nut stores that the parent birds accumulate in the autumn. How is this regulation of the clutch size achieved ? The hazel nut stores are used by the adult birds both for their own consumption in winter and as an important food source for the raising of young in the following spring. Assume that the stores were utilised for adult bird consumption at a rate that was similar each winter. After a poor hazel nut year when the total stores were small, the stores would be dependent before egg laying started an no nuts would be available for the young. In other years stores might last until egg laying but not until hatching. Under such conditions the birds´ lipid and protein reserves could be high and the birds lack a reliable cue for " setting the numerostat". Both 3-egg and 4-egg clutches would be likely to be laid and nestling starvation would occur. However, Nutcrackers usually have nuts available for the young after an autumn with a poor hazel crop. This strongly suggest that they, in winters when stores are small or average, regulate their own use of the stores in a way that normally ensures sufficient nuts remaining available for the young. The Nutcracker has the capacity to remember not only the exact location of each nut cache but also which cache have already been used earlier in the winter. This capacity is so great that, after an autumn with a very rich hazel nut crop when some caches remain unused over the following summer, the bird accurately locates caches that were made 17 months earlier. It is conceivable that a bird with such a "mental power" might be able to regulate the use of its winter stores in accordance to their size.
The upper limit of the clutch size may be influenced by the birds´ ability to provide proper incubation temperature to the eggs. The weight of the fresh 4-egg clutches of Nutcracker A and B were 24 and 23.4 % of the weight of females registered 5 to 6 weeks after laying. The author once recorded egg laying when ambient temperature was 55 degrees C below adequate incubation temperature. For the Slender-billed Nutcracker N.c. macrorhyncos in Siberia the difference between ambient and incubation temperatures may be even larger.
Only a few adequately checked clutches of N.c. macrorhyncos have been mentioned in literature. Two clutches consisting of three eggs were found by Reimer (1959) and one of 4 eggs by Karpuchin (1962). One 3-egg and two 4-egg clutches are to be found in egg collections in Sweden.Thus the clutch size in N.c. macrorhyncos may be the same as in N.c. caryocatactes.
Two factors regulate the population of the Thick-billed Nutcracker in Scandinavia:
Access to a sufficient amount of hazels within reach of the nesting habitat.
Conifer forest of such quality and extent that it fulfils the demand on a territory, i. e. it serves as a safe caching area and allows successful breeding.
Table 2 shows the size of clutches in Swedish egg collections; the relation between clutch size and hazel crop is much less clear than in tab 1. It is not known why this is so, but it illustrates the words by David E Davis (1955) "Rarely are data from miscellaneous sources adequately controlled to serve special purposes".
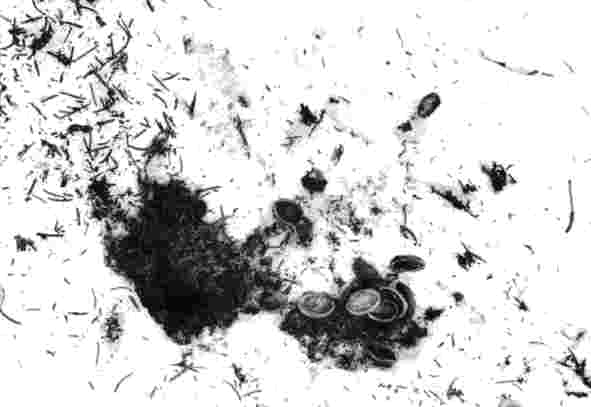
Ruvfläckar hos nötkråkan är blott några exempel på de fynd som P O Swanberg gjorde under sina många och långa tider tillsammans med kråkorna på Billingen.
Nötkråkans Nucifraga c. Caryocatactes vana att lägga upp vinterförråd
![]() The hoarding habit of the Nutcracker (Nucifraga c. caryocatactes)
The hoarding habit of the Nutcracker (Nucifraga c. caryocatactes)
Per Olof Swanberg
Att den tjocknäbbade nötkråkan, Nucifraga c. caryocatactes, samlar nötter om hösten, visste redan Carl von Linné för mer än två hundra år sedan. Det är förvisso förlåtligt, att den annars häpnadsväckande skarpsynte Linné dock i sina föreläsningar 1748-1752 yttrade, att visserligen "gömmer hon" nötter, men "eftersom hon tar märke på molnen, som bonden säger, så finner hon ock intet igen sin skatt, då märket förswunnit".
Ännu för fyrtio år sedan var nötkråkans biologi föga känd. När jag kommit till Skara 1935 och funnit, att det fanns en fast population av arten på Billingen, blev det därför naturligt att intensivstudera den. Det var bl.a. två sidor av biologin, som befanns vara mycket fängslande objekt för ingående studier. Den ena var vanan att bygga bo och lägga ägg redan i vinterkyla i mars - när jag en gäng, den 17 april 1938, skulle visa mitt första nötkråkbo för en grupp från Dansk Ornithologisk Forening fick vi plumsa fram genom snö till knäna, när nötkråkan på sitt bo med små ungar i en låg gran långt inne i granskogen på platån ovan Öglunda skulle beskådas. Den andra märkliga egenskapen hos fågeln blottades undan för undan: det stod så småningom klart, att fågeln inte bara systematiskt lägger upp vinterförråd av hasselnötter i sitt häckningsrevir och flitigt utnyttjar dessa otaliga, utspridda småförråd utan också har förmågan att återfinna dem under snötäcke.
Så småningom stod det klart, att nöt hamstringen och förmågan att utnyttja de upplagda vinterförråden är helt livsavgörande för nötkråkan. För att få fram ett mått på fågelns förmåga att återfinna sina vinterförråd gick jag till väga på det sättet, att jag varje söndag (lördagar hade man inte fria på den tiden) skidade kors och tvärs i nötkråkrevir, sökte upp de gropar i snön, genom vilka nötkråkan sökte sina nötter, och letade upp var i närheten hon hade knäckt de återfunna nöttema. När jag sammanfattade resultatet av 351 funna gropar i snön befanns, att nötkråkan bevisligen i 86 % av fallen funnit sina nötter och att hon prickat precis rätt, när hon grävde sig ned genom snötäcket, hur djupt detta än var.
Ännu märkligare: dessa fynd fördelade sig med en tredjedel på vardera av månaderna januari, februari och mars. Det befanns, att procenttalet "fullträffar" var exakt detsamma för alla tre månaderna. Detta betyder, att nötkråkan inte bara har en nästan obegriplig förmåga att minnas och trots kanske halvmeterdjupt snötäcke lokalisera varje liten gömma i mossan (alla förråd göms en näbblängd djupt i mossan och täcks noga) utan hon har också ett abstrakt minne, som i t.ex. mars talar om för henne vilka förråd hon tömt i december-januari, så att hon aldrig letar två gånger på samma punkt.
Märkligt är också, att hon om hon efter ett extremt rikt nötår har kvar outnyttjade förråd, ännu efter 16-17 månader, när det goda nötåret kanske följts av ett dåligt, kan leta upp sådana förråd, som då legat i mossan genom två vintrar. Det är lätt att på skalen se, att de legat två säsonger i mossan. Nötkråkan minns alltså fortfarande, vilka förråd hon inte utnyttjat under en föregående vinter.
I själva verket är det rimligt att antaga, att träffsäkerheten är högre än 86 %, kanske är den nära 100 %, ty dels får man räkna med brister i den mänskliga observationsförmågan, dels får man räkna med att ett och annat förråd kan ha påträffats och tömts av skogsmöss.
En enda gång har jag tydligt fått bevittna själva gömmandet av ett reguljärt vinterförråd. Det var den 11 september 1949 i den glesa tallskogen på ön Sottunga i Ålands skärgård. Jag satt i en trädtopp för att registrera nötkråkornas revir på ön och kunde då bevittna hur en nötkråka kom med en börda nötter från en hasselbacke och tömde och gömde den intill några långa gula grässtrån på mossklädd hällmark. Efteråt gick jag dit. Inget tecken visade, att där kunde finnas något gömt, men när jag lyfte på ett löst täcke av mossa, blottades ett förråd av 18 nötter, väl packade och staplade två skikt precis som bitarna i en sillburk. Antalet, 18 st., torde vara ungefär normalt för en börda av våra små nordiska hasselnötter. En av mina meddelare, Karl Larsson i Skara, omtalade 1942, att han räknat 18 nötter i "krävan", och Tore Hansson i Örebro meddelade 1943, att han en gång under något tidigare år funnit 21 nötter i en nötkråkas hakpåse.
Nötterna befrias alltid från sitt hylle, innan de stoppas ned i det speciella transportorganet, som är en sublingual (under tungan) säck eller hakpåse, vilken, som namnet säger, mynnar med en öppning under tungan. Det gamla bruket i olika länder att kalla detta organ "strupsäck" är alltså inte riktigt adekvat, vilket har påpekats av tre amerikanska anatomer, W. J. Bock, R. P. Balda & S. B. Vander Wall, i tidskriften The Auk 1973. Denna speciella nötpåse stoppas som bekant så full, att man med blotta ögat på 70-90 meters håll och med en god kikare på nära kilometerhåll kan se den under hakan oformligt utspända, nedhängande "påsen". Endast i något mycket sällsynt undantagsfall transporterar en nötkråka en hel nötklase i näbben under den långa hamstringsfärden. En och annan gång kan hon dock under färden hålla en nöt i näbben. Detta är emellertid undantagsfall till skillnad frän nötskrikans, Garrulus glandarius, beteende - den sistnämnda bär som bekant ofta ett ollon väl synligt i näbben när hon är sysselsatt med sina insamlingar.
Nötkråkans häckningsrevir ligger - sedan miljoner år kan man med gott samvete säga - alltid i barrskog. Eftersom vår svenska ras, den tjocknäbbade, är specialinriktad på hasselnötter, får Billingens nötkråkor normalt flyga 2 - 6 kilometer mellan insamlingsplatsen och häckningsreviret uppe på platån. Alla vinterförråd lägges nämligen inom revirets gränser, dock så att varje ny börda göms på en ny punkt i marken i reviret. De nötkråkor, som varje höst passerar mitt hus i Bjällum under hamstringsturerna från hasselbackarna i Bolum, har 3-4 km att flyga, och Barbro & Thore har bevittnat, hur nötkråkor, som samlat hasselnötter på Ore backar vid Dagsnäs, försvunnit med sådan färdriktning, att de som slutmål måste ha haft revir på Billingen ca 10 km från startplatsen.
Nötkråkan tillbringar hela vintern i sitt 12-15 hektar stora revir i djup granskog. Hon lägger inte upp vinterförråd av annat än nötter - hon är alltså mer ensidigt specialiserad än nötskrikan, som gömmer även annat än ekollon. Det är alltså viktigt för henne att det är "prima vara" hon gömmer. Nötterna skall räcka både till henne själv, när det är minus 30 grader i skogen, och till föda åt ungarna i boet, i synnerhet när skogen är snötäckt. Det är därför intressant att konstatera, att hon aldrig hamstrar tomma nötter och ännu märkligare, aldrig nötter som är angripna av nötviveln!
För att få en uppfattning om hur hon avgör, om en nöt är tom, gjorde jag följande experiment: ett antal kraftiga nötter utvaldes, klövs försiktigt, kärnan avlägsnades och skalhalvorna limmades samman så noga, att skarven ej skulle märkas. Sedan utvaldes samma antal välfyllda mindre nötter med exakt samma vikt som de tömda. Alla lades ut till mina snälla studie- nötkråkor, som helt accepterat mig i sitt revir. De plockade ivrigt åt sig de fyllda nötterna, men när de tog en tom nöt av samma vikt i näbben, kastades den ögonblickligen så kvickt som om den brändes. Det var alltså inte själva vikten, som var avgörande, utan densiteten eller möjligen resonansen i den tomma nöten.
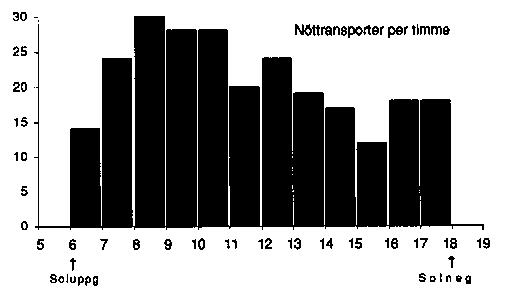
Fig. 1. Diagrammet visar antalet nötkråkor som med hakpåsen fylld med nötter sågs passera observationspunkten i revir A den 26 september 1948.
![]() Number of flights of Nutcrackers carrying nuts in the sublingual pouch, observed from a
fixed point in territory A, Sept. 26,1948.
Number of flights of Nutcrackers carrying nuts in the sublingual pouch, observed from a
fixed point in territory A, Sept. 26,1948.
Nötkråkan har en för oss människor fabulös förmåga att lokalisera sina "otaliga" förråd, även om halvmeterdjupt snötäcke suddat ut nästan allt som i vårt tycke kunnat tjäna som märke för minnet. Hon gräver sig inte lodrätt ned genom snön utan snett ned, och gången genom snön är inte vidare än vad som behövs för hennes kropp och slutar ändå exakt där nöttena stuckits ned. Tydligtvis vägledes hon av något som kan kallas muskelminne, dvs. hon har i minnet registrerat exakt hur hon rörde sig, när hon valt punkten för nötgömman. För att pröva henne har jag i hennes åsyn i reviret presenterat nötter och gömt dem i mossan bara 8 - 10 meter från hennes uppmärksamma blickar. Nötkråkan har sedan, när jag tagit några steg tillbaka, hoppat ned och letat men inte kunnat finna nöttena, trots att hon åsett precis var jag gömde dem på hennes eget maner. "Felet" var alltså, att hon inte gömt dem med sin egen rörelseapparat! Naturligtvis kan en nötkråkforskare inte underlåta att undersöka kvalitén hos nötter förvarade à la nötkråka. Jag gömde alltså färska nötter precis som nötkråkan i mossa uppe i granskogen på Billingens platå, dock försiktigtvis under finmaskigt metallnät. Ännu på eftervintern smakade dessa nötter som färska, nymogna. Jag rekommenderar alltså metoden för dem som vill ha utsökt smakande julnötter.
I detta sammanhang kan det kanske vara värt att nämna ett annat exempel på vinterförråd i naturen. Efter det att jag upprepade gånger i nötkråkskogen kunnat studera, hur skogsmården lokaliserat sina gömmor och grävt upp tjäderägg i mars månad, sannolikt omkring tiden för ungarnas födelse, gjorde jag det experimentet, att även jag gömde ägg i mossan på det sätt, jag lärt av mården, dock med den skillnaden, att jag tog hönsägg från det egna skafferiet. Vid ett tillfälle, då Göteborgs ornitologer kom hit för att se nötkråkor i mars, grävde jag sedan upp mina ägg i vännernas närvaro. Äggen var friska, även om min familj lät mig konsumera dem ensam, sedan de stekts i köket.
För att kunna visa hur insamlingen av nötter totalt dominerar för att inte säga fyller nötkråkans tillvaro, så länge nötter finns att tillgå, användes följande metod. I ett nötkråkrevir "A" utvaldes en av de högsta granarna så högt belägen, att man från dess topp kunde överblicka all granskog på minst en kilometers avstånd åt olika håll. I denna fastsurrades en fotbräda så högt som möjligt, och sedan kunde jag vid observationstillfällena, stående på brädan, surra fast mig själv med en specialgjord livförsäkringsrem. Fickuret hängdes på toppspiran framför ögonen, den specialgjorda kollegiebladboxen hängdes i snabb-beredskap och på sådant sätt hade jag hela tiden båda händerna fria för kikaren. Även maten måste placeras snabbåtkomlig inom räckhåll, innan dagens aktivitet inleddes.
Från denna observationspunkt kunde nötkråkornas insamlingsfärder inom ett avstånd av nära en kilometer noggrant registreras. När nötkråkan kommit in över den kompakta granskogen på platån, flyger hon nämligen konsekvent i praktiskt taget rak linje lågt över grantopparna utan uppehåll direkt till reviret. Först där dyker hon ned bland träden.
1948 var ett rikt nötår. I slutet av september, när nötterna spontant lossnade ur klasarna, nådde hamstringen sin höjdpunkt, och den 26 september registrerades alla hamstringsfärder, som under dagens aktiva tid passerade min observationspost. I allt noterades nötkråkor med fylld hakpåse passera 252 gånger, och verkliga antalet bör ha varit något litet större, dels därför att uppflygande nötkråkor hade motvind och därför flög ovanligt lågt över trädtopparna och i vissa fall kunde vara vanskliga att upptäcka, dels enligt följande anteckning mellan kl. 17.30 och 18.10: "I verkligheten borde här varit flera, ty nu är det så mörkt (mot mörk himmel dessutom!) att jag ej torde upptäcka perifera nötkråkor". 38 gånger anlände någon av nötkråkorna med sin börda till det revir A, där jag posterat mig. Räknar man hypotetiskt med ca 18 nötter per börda, blir summan för ett enda par under en enda toppdag alltså ca 700 nötter. Sannerligen ett målmedvetet, oförtrutet arbete för familjens framtida uppehälle.
Av det nyss sagda framgår, att om man skall söka nötkråkor i ett visst område, är de första tre veckorna av september gynnsammast: då far nötkråkorna normalt från morgon till kväll i rastlös trafik mellan hasselsnåren och häckningstillhållet i barrskogen.
Eftersom 1948 var ett rikt nötår, blev det även ytterst lämpligt att registrera, hur hamstringssträcket fördelar sig över hela säsongen. Sträcket satte i gång vid mitten av augusti och nådde sin höjdpunkt i senare delen av september. Den 5 december kom mina två A kråkor med bara tre bördor nötter. Den sista av dem kom redan kl. 12.07 nedifrån sluttningen och hade då ingen börda nötter med – detta blev slutsignalen, som visade, att årets insamling avslutats: när jag den 19 december i klar sikt hela förmiddagen stod i min granspira, syntes alltså inte en enda nötkråka passera. De stannade nu i reviret. Observera dock att detta var under ett mycket rikt nötår. Under år med mindre nöttillgång slutar givetvis trafiken tidigare, ibland mycket tidigare.
Denna observationsmetod är utmärkt även för noggrann registrering av en populations revir. Under åren 1944-1946 gjorde jag med den metoden en totalinventering av alla revir på centrala och norra Billingen norr om landsvägen Skövde - Varnhem. Resultat: 145 nötkråkepar norr om landsvägen på Billingen, det vill säga ca 4 par per km2. Stickprov söder om landsvägen gav anledning att skatta totalpopulationen på hela Billingen till omkring 200 par. Till jämförelse kan nämnas, att reviren på ön Sottunga utanför Åland kunde inventeras på 3 dagar, 8 - 11 september 1949, med resultat: 40 par på 360 ha skogsyta. Den ringa revirstorleken där, ca 9 ha, kan sättas i samband med flera faktorer: omgivande vatten tenderar att tränga ihop nötkråkorna, som inte tycker om att flyga ut över vatten, det maritima klimatet ger jämnare fruktsättning hos hasseln och, inte minst avsaknaden av konkurrerande- parasiterande smågnagare på ön.
Det säger sig självt, att en så högspecialiserad egenskap som nötkråkans förmåga att utnyttja vinterförråd inte skulle kunna utbildas som karakteristisk för arten, om den inte hade livsavgörande betydelse. Ovan har nämnts, att nötter utgör den viktigaste födan under vintern och är oundgänglig även som föda åt ungarna i boet. Detta fick jag belägg för efter det katastrofalt dåliga nötåret 1943. Följande år hade nötkråkorna i reviret B det normala minimiantalet ägg, 3 st., som kläcktes på morgonen den 1 maj. Den 17 maj antecknades om ungarna: "upptar bara hälften av boet. Normalt borde de ju nästan fylla det. Ser små och magra ut. Tydligen panikartat hungriga, ty redan när jag började klättra vid basen av stammen, började en väldig konsert av kvillrande pip.." Det sistnämnda var något unikt, ty normalt är nötkråkungar alltid tysta i boet, även när föräldrarna kommit dit. Den 22 maj antecknades, att "ungarna se ut som små dvärgar" och att det gått så långt, att de nu skrek även när de var ensamma i boet och detta så ljudligt att det hördes tydligt på 160 meters håll. En unge låg redan död i redet, och vid en kontroll två dagar senare var detta tomt, ungarna tydligen ihjälsvultna. Eftersom nötter är stapelföda för ungarna under deras tid i boet och eftersom föräldrarna ej haft möjlighet att lägga upp nämnvärt vinterförråd av nötter, svalt ungarna alltså ihjäl.
Hösten 1943 hade vi av samma orsak haft den största dittills kända utvandringen av tjocknäbbade nötkråkor. De gamla exemplaren med etablerade revir på Billingen stannade kvar, men de unga, som i augusti hade släppt familjebanden och skulle söka upp sådan nöttillgång, att de kunde samla vinterförråd för egen etablering, blev tvingade att vandra ut. Det är tydligt, att utvandringen kan sägas vara ett förlängt hamstringssträck, utlöst av att inget finns att insamla för vinterförråd. Ty allmänt sett finns det ännu gott om föda åt fåglarna själva i augusti, när sträcket plötsligt startar.
Hösten 1975 fick vi bevittna en upprepning av företeelserna från 1943. När jag gjorde min årliga hassel- fruktsättningskontroll den 29 augusti, fann jag 9 nötter på sammanlagt 91 buskar mellan Skara och Billingen, alltså en nöt på 10 buskar. När jag gick tillbaka till anteckningarna från 1943, sågs nästan identiskt samma situation: "23/8 1943. Var tionde eller tjugonde buske har 1-2 nötter": Det är lätt att inse, att detta är en katastrofal brist jämfört med normala år. För att illustrera skillnaden med siffror kan nämnas, att min noggranne fader gav mig följande tal från sina 60 buskar i Nyvång i Skåne: 1943 skördades summa 32 nötter, år 1945 31 kg, det vill säga ca 14 400 nötter, alltså en skillnad motsvarande ungefär 1/450.
Den 17 och 24 augusti 1975 kunde man se nötkråkor flyga med bördor av nötter på Billingen. Men redan den 30 augusti var den trafiken slut. Det blev som väntat en stor utvandring av svenska nötkråkor.
Eftersom nöttillgången i landet hösten 1974 varit ymnig, måste nötkråkornas reproduktionsresultat 1975 ha varit mycket gott, vilket ökat utvandringens numerär. Gunnar R (1975) meddelar också "att rasen caryocatactes aldrig tidigare uppträtt vid Falsterbo i ens tillnärmelsevis liknande antal". Å andra sidan kan man förstå, att reproduktionsresultatet 1976 liksom 1944 kommer att bli under det normala, särskilt accentuerat om det skulle bli en sen vår.
Ruvning hos nötkråkan
Ruvning hos nötkråkan, Nucifraga c. caryocatactes publicerat i Grus Skövde fågelklubbs tidskrift.
Nötkråkan häckar som bekant mycket tidigt på året. Äggen lägges vanligen i sista tredjedelen av mars eller första veckan av april. Det innebär att det kan bli mycket kallt under tiden, långt under -10 grader. Det är alltså naturligt, att nötkråkan ligger på boet redan från och med första ägget.
Mannen deltar i ruvningen
I hela den stora ordningen Passeriformes finns bara ett fåtal undantag från regeln att hanen icke deltar i ruvningen av äggen. Vanligt är, att hanen hos olika arter i stället matar den ruvande honan. Det fungerar bra även hos så tidiga häckare som t.ex. korsnäbbarna, Loxia, som rättar sin häckningstidpunkt efter den period, då barrträdskottarna börjat släppa sina frön och hanen alltså direkt från källan kan skörda vad som behövs för det dagliga behovet.
Nötkråkorna har andra villkor: de kan visserligen tillgodogöra sig granfrö, när kottarna kränger, men deras specialföda under hela häckningssäsongen är hasselnötter eller de i hårda skal inkapslade frön av cembratallen, som samlas av den smalnäbbade nötkråkan i Sibiriens taiga. I Schweiz samlar de tjocknäbbade nötkråkorna i första hand cembrafrön men även, särskilt i Jurabergen, hasselnötter.

Figur 1 Honan avlöser hanen vid bo B den 8 maj 1952.
The Female B relieves the male.
Förråden skall trygga även uppfödningen av ungarna i en period, då marken är snötäckt och nästan ingen annan mat finns att säkert tillgå. Förråden är individuellt upplagda, och hanen vet inte, var honan gömt sina nötter (Swanberg 1976). Det är alltså naturligt, att var och en av dem måste hålla reda på och utnyttja sitt eget nötförråd. Konsekvensen blir, att hanen måste skydda eller ruva äggen, när honan är ute i snön. Detsamma gäller för den amerikanska nötkråkan, Nucifraga columbiana. L R Mewaldt (1956:16) konstaterade vid ett bo av denna, att hanen tog 20 % av ruvningstiden mellan kl. 0715 och 1800.
Undersökningar 1938 - 1952 i granskogen på Billingens platå visade klart, att hane och hona av tjocknäbbad nötkråka delar ruvningsbestyren och gör detta efter i stort sett sådant mönster, som framgår av diagrammen fig. 2. Det innebär, att honan ruvar under natten och avlöses av hanen strax före eller omedelbart efter soluppgången. Avlösningen går alltid sekundsnabbt, honan lämnar inte plats, förrän hanen sitter på bokanten. Hanens första ruvningspass är alltid tämligen kort, 20 - 80 minuter, därefter tar han ytterligare två eller tre ruvningspass under dagen, men honan får börja kvällens och nattens långa pass redan flera timmar före solnedgången.

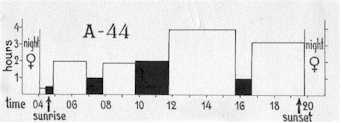
Ruvningens fördelning mellan hane och hona hos två par nötkråkor. A 44
under sjunde dagen av aktiv ruvning och B 52 under femte ruvningsdagen.
Shares in incubation: A 44 on the 7th day of active incubation, B 52 on the 5th
day.
Fig 2 åskådliggör att under den tid på dygnet då nötkråkor är aktiva, ruvande hanen A 26% av tiden , hanen B 43 %, medianvärdet blir alltså 34%. Detta kan kompletteras med resultatet av 68 observationstimmar under 13 olika dagar vid 7 andra bon. Dessa gav resultatet, att hanen ruvade genomsnittligt 46% av observationstiden. Emellertid bör observeras, att observationerna vid dessa 7 bon utfördes före kl. 12. Jämför man med nötkråkorna A och B i diagrammet, kommer man fram till att även hos dem tog hanen i medeltal 48% av ruvningen före kl. 12. Överensstämmelsen är alltså god och ger anledning att betrakta de nämnda diagrammen som signifikativa för arten, åtminstone i Sverige. Skillnaden mellan förmiddag och eftermiddag ifråga om makarnas arbetsfördelning är ännu en påminnelse om att dagsrytm hos en fågel inte utan vidare kan utläsas ur summan och medelvärdet av slumpvis gjorda korttids observationer.
Ruvfläckar
Länge trodde man, att "ingen hane av Passeres ruvar" (K. Kolthoff I934:55, se även Bailey 1952). Följaktligen ansåg man då, att inga hanar av Passeres har ruvfläckar. Kjell Kolthoff konstaterade visserligen, att han ej sällan funnit, att hanen hos en del tättingar kan visa naken undersida, men tillägger, att huden då ej företer sådana vävnadsförändringar, som utmärker äkta ruvfläckar. Troligen är det motsvarande företeelse, som Becker & Ringleben (1975) funnit hos citronsiska, Serinus citrinella, och kallar "pseudo-ruvfläck". Beträffande just kråkfågel kan nämnas, att M.J. Erpino (1969:269) fann, att en av tre hanar av skatan, Pica pica, som undersökts, sedan honorna lagt ägg, visserligen på buksidan visade helt naket apterium men att huden där i sin struktur ej ombildats till ruvfläck (apterium= hudparti utan fästen för konturfjädrar men normalt dunklätt).
Hos stare, Sturnus vulgaris, vars hane deltar i ruvningen, fann dock J.A. Lloyd (1965) utbildad ruvfläck.
Med tanke på hanens roll vid äggens ruvning hos nötkråkan och överensstämmelse i ruvningstidens längd hos olika par kunde man dock dra slutsatsen, att det vore otroligt, att hanen hos denna art ej skulle ha utbildade ruvfläckar, som fungerade lika väl som honans. För den skull fångades sex hanar av nötkråka under den tid, de hade ägg eller ungar i bo. Alla sex visade en ruvfläck av samma storlek och alldeles samma makroskopiska, för ruvfläckar specifika struktur som honornas vid samma tid.

Fig 3. Hane B visar på denna bild ungefär den ställning som fåglarna intar när de sitter på boet för att lämna endast skyddande värme, ej ruvningsvärme.
The male B, in brooding position, corresponding to its behaviour when it is brooding the eggs before true incubation.
Ruvfläcken omfattar hela undersidans apterim från framkanten av sternum, bröstbenet, till kloaköppningen under stjärtroten. Storleken framgår av följande två exempel: hane B fångad den 17 maj 1953, 25 dagar efter sista äggets kläckning visade följande mått på ruvfläcken: längd 76 mm, bredd över bröstet 45, över buken 37 m.m. Hona B som vår något mindre än hanen, fångades den 3 maj och visade därvid följande mått på ruvfläcken längd 70 mm, bredd över bröstet 50, över buken 33 mm.
För att studera utvecklingen av ruvfläcken i förhållande till äggläggningen fångades makarna i paret D den 21 mars 1954; d.v.s. 1 - 2 dagar före bobyggnadens början eller 12 dagar före första äggets läggning. Det visade sig, att hanen då, 14 dagar innan kullen skulle vara fullagd, hade tappat dunet på blott 25% av den kommande ruvfläckens yta. Hos honan var bara en kvcm hud naken, alltså ca 5% av den färdiga ruvfläcken. Redan 7 dagar senare, alltså 5 dagar före första ägget, befanns att både hanens och honans ruvfläck nått normal längd och nästan normal bredd, men att huden ännu ej erhållit den hyperplastiska struktur och den blodförsörjning genom vidgade kärl, som senare skulle följa.
Därmed lämnades de snälla studiekråkorna D tacksamt i fred, och den 11 april undersöktes i stället honan B under den dag, då hon lagt sitt sista ägg i årets kull. Hennes ruvfläck var då fullt utbildad med huden lös, starkt skrynklad och rikt blodförsörjd. Skillnaden i hudens vävnadsstruktur, då den utbildas till ruvfläck, framgår bl. a. av att motsvarande hudparti hos skatan före häckningstiden väger 1 gram, medan samma parti efter äggens läggning väger 7 gram (Erpino 1969).
Den viktiga mikroskopiska strukturen hos olika arters ruvfläckar, bl. a. hos kråka och råka, är ingående beskriven av Lange (1928: 689-693) och ruvfläckens mikroskopiska anatomi hos Nucifraga columbiana har beskrivits av Bailey (1952), och även hos den arten har hanen en väl utbildad ruvfläck (Mewaldt 1952:361). Själva utbildningsförloppet har hos skatan specialstuderats av M.J. Erpino, och hos den kaliforniska tofsvakteln, Lophortyx californticus, av R E Jones (1971 med rik litteraturförteckning).
Ruvningsteknik
Trots att nötkråkan stadigt ligger på boet alltifrån första ägget kläcks ungarna normalt under loppet av en och samma dag. Kortaste kläckningsperioden för 15 kontrollerade kullar noterades till max 6 timmar (3 äggkull B 44), en längsta till ca 24 timmar. Länge spekulerade man över förklaringen till att alla ungar kläcks ungefär samtidigt även hos en del sådana fågelarter, som tycktes ruva från första ägget. En del författare tänkte sig, att utvecklingstiden för de först lagda äggen kunde vara längre än för senare ägg i samma kull. De första som kom med en godtagbar teori var Ryves (1929) och Holstein (1943, 1950). De antog, att den temperatur, som fåglarna ger de första äggen, ej räcker för att starta och underhålla utvecklingen av ett embryo.
Detta föreföll vara den riktiga förklaringen. Att visa det blev en intressant uppgift. Jag började med att förfärdiga ett naturtroget målat plastägg, försåg detta med termoelement, förbundet med ledning genom boets botten till en urverksdriven skrivare under boträdet. Resultatet blev ej tillförlitligt, och metoden var för komplicerad och störande för arbete med flera olika bon i djupet av den väglösa storskogen.
För genomgående kontroll av så många kullar som möjligt användes i stället först metoden att approximativt undersöka temperaturen hos äggen genom att trycka dem mot den känsliga överläppen. För kontrollerad gradering av förnimmelsen hade jag medfört hönsägg, som gavs olika, med kvicksilvertermometer registrerad temperatur och i nötkråkereviret prövades mot läppen. Denna metod tillät en nöjaktigt god uppfattning av temperaturen vid det snabba arbete, som var nödvändigt vid den låga lufttemperaturen, ofta under fryspunkten.
Det gick att på detta sätt grovt uppskatta temperaturer, mellan, 20° och 35°. Effektiv ruvningstemperatur hos fåglar anses ligga mellan 33° och 40° (Huggins 1941), med 34° som vanlig temp bland Passeriformes (Tucker 1943) och Stresemann (1934) uppgav, att ännu vid 28° äger ingen utveckling rum. Följande fynd må ses mot detta bakgrund.
Temperaturen hos det första ägget under den dag, då detta lagts, kontrollerades i 22 bon. 13 av dessa var äggen kalla, trots att fågeln låg på boet. I övriga 19 fall noterades, att äggets ovansida hade en temperatur mellan 20 och 28°, genomsnittligt ca 22°.
Den dag, då andra ägget lades, befanns äggen i 4 bon vara kalla, trots att fågeln låg på boet. I 27 fall skattades temperaturen till genomsnittligt 23 grader. I endast ett fall föreföll temperaturen något överstiga 30°.
Den dag, då tredje ägget lades i 4 äggkullar, skattades äggtemperaturen i 17 bon till i medeltal 26'.
När kullen blivit fullagd, noterades alltid, att äggen dels var jämnvarma runt om, dels höll en temperarur som beräknades ligga kring 35' C.
Detta indikerade, att hos nötkråkan ingen eller obetydlig embryonalutveckling äger rum under äggläggningstiden trots att fågeln hela tiden ligger på boet, och det visar alltså, att full ruvningsvärme ges åt äggen först sedan kullen blivit fullagd.
Ruvningstid
För att bevisa, att utvecklingstiden för embryot i alla äggen i kullen är ungefär densamma, gjordes nedannämnda experiment, vilka praktiskt underlättades av att nötkråkans ägg befunnits läggas med något mer än 24 timmars intervall. Se tabell 1 nedan.
| Tid på dagen för läggning av kullens olika
ägg |
||||
| Ägg nr Egg no. |
Antal Number |
Äggläggning ej funnen Egg laying not proved |
Antagen normal läggningstidpunkt supposed most frequent time of laying the egg. |
|
| 1 | 18 | 0600 | 0900 | 0530 - 0800 |
| 2 | 17 | 0715 | 0800 | 0700 - 0800 |
| 3 | 17 | 0750 | 1300 | 0800 - 1200 |
| 4 | 9 | 1100 | 1422 | 1100 - 1400 |
Tabell 1
Experimenten gick principiellt ut på att låta nötkråkor med i full gång varande ruvning av färdiglagd kull ta hand om ruvningen av första ägg från nötkråkor med några dagar senare äggläggningstid. Det gällde alltså att noga hålla reda på förehavandena i flera, till varandra gränsande nötkråkrevir. Arbetsschemat var detta:
- 1. Nylagda ägg nr 1 eller 2 flyttades snabbt över till annat bo med i full gång varande ruvning. (För att fågeln ej skulle reagera negativt, lades ett rätt tempererat imiterat ägg i det avlägsnade äggets ställe.)
- 2. Sista ägget i vissa kullar flyttades omedelbart efter läggningen över till ett bo, där första ägget lagts något tidigare samma dag.
| Behandling av ägg nr 1 |
Egentlig ruvningstid True development time, days and hours |
| B1-47. Lagt 8/4 före 08.00, flyttat till A-47 09.00, där sista ägget lagts föregående dag 14.22. Kläcktes hos A-47 26/4 f.m. eller mitt på dagen. | 18 dygn + 0-10 tim |
| B2-47. Lagt 2114, utbytt mot ett ägg nr 4 kl 18.20. Förvarat i rumstemp, fört 24/4 06.10 till A-47, som då ruvade aktivt. Tillbaka till 82-47 26/4 18.55. Kläcktes 12/S 04.20, samma morgon som övriga kullsyskon. | 17 dygn + 22 tim |
| B-48. Lagt 3/4 före 10.00. Flyttat till A kl 10.30 (3 dagar efter läggn av ägg a nr 4). Flytta: tillbaka till B 10/4 18.00. Kläcktes 21/4 före 18.00, d.v.s. 3 dagar före kullsyskonen ägg nr 3 och 4 i samma bo | 18 dygn + 0-7 tim |
| B-50. Lagt 2/4 före 09.20. Flyttat till E 09.50 (dagen efter läggn av ägg nr 4 i E). Kläcktes på morgonen 20/4, 1/2-1 dag senare än E nr 1-3. | c 18 dygn |
|
Tab 2. Utvecklingstid för ägg nr 1, sedan det flyttats till bo, där aktiv ruvning redan startats med en annan nötkråkas fjärde ägg. - De angivna värdena skall jämföras med det förhållandet, att ägg nr 1 i 4-kullar normalt kläckes 18 dygn + 71,5 tim efter dess läggning, medelvärde, medan medelvärdet för ägg nr 4 är 18 dygn minus 1 timme. (Anm. motsvarande tabell för ägg nr 2 finns hos Swanberg 1956). Development time of egg no 1 after it had been placed in a nest where another nutcracker had 4 eggs and active incubation consequently was in progress. - The noted figures may be compared with the fact, that egg no 1 in a 4-egg-cIutch normally on an average is hatched 18 days + 71,5 hours after its deposition, whereas the mean figure of eggs no 4 was found to be one hour less than 18 days. (Corresponding figures of egg no 2, cf Swanberg 1956). |
|
tabell 2
Resultaten av dessa förflyttningar framgår av några exempel i tabell 2 och visar klart, att utvecklingstiden för embryot i de olika äggen är fysiologiskt lika i vilken ordningsföljd än respektive ägg har lagts. Den verkliga eller effektiva ruvningstiden är alltså densamma för alla äggen i kullen.
En viss individualitet finner man dock, ej oväntat, hos de enskilda äggen i utvecklingstid liksom även i storlek och form. Sålunda hände bara i 2 fall av 13 i detta hänseende tillräckligt noggrant kontrollerade kullar, att alla äggen kläcktes precis i den ordning de lagts. I boet A-45 kläcktes ägg nr 4 före de tre andra. Men i 10 av 12 kullar kläcktes det sist lagda ägget sist. Detta visar, att en viss embryonalutveckling verkligen kan börja före kullens sist lagda ägg, samtidigt som det, tillsammans med övriga redovisade data visar att denna utveckling i sådana fall går så långsamt, att skillnaden i de olika äggens kläckningstidpunkt endast är ett fåtal timmar.
Av det föregående framgår nötkråkans ruvningstid. Emellertid har mycken oklarhet i äldre litteratur orsakats av olika uppfattning om hur artens ruvningstid skall anges, då fåglarna iakttages ligga på boet redan under äggläggningsperioden. Äggets ruvningstid är visserligen den tid, som det ruvas med full ruvningsvärme - men hur fastställa ruvningsvärmens början för det enskilda ägget? Därför må nämnas, att man nu, av praktiska skäl, med en arts ruvningstid menar tiden från läggning av sista ägget till dettas kläckning (Nice 1954, Swanberg 1956) och att man om sista ägget ej blivit numrerat, kan acceptera att använda tiden mellan läggning av sista ägget och kläckningen av sista ungen i kullen (Swanberg 1950).
Vid beräkningen av ruvningstiden i fält kan man i regel inte exakt på timmen fastställa läggningstid och kläckningstid. Det skulle bli alltför störande för fågeln. Man är alltså vanligen hänvisad till att ange tiden till mellan två värden, t.ex. mellan 17 dygn + 23 timmar och 18 dygn + 2 timmar, och sedan ta medelvärdet för ett antal kullar.

Fig 4. Honan B nedtryckt i boet, vars inre djup är 8,5-9 cm. under sextonde dagen av aktiv ruvning.
The female B sitting in typical incubation posture, suppressed in the 8,5-9 cm deep nest cup, photographed in the 16th day of incubation.
För nötkråkans del erhölls följande värden för 17 kontrollerade kullar.
3-äggkullar (4 st.): medelvärde 18,16 dygn.
4-äggkullar (13 st.): medelvärde 18,0 dygn. Det är alltså berättigat att kort och gott säga, att ruvningstiden är 18 dygn. Som jämförelse må nämnas, att Medwaldt (1956) i två bon av den amerikanska nötkråkan också fann ruvningstiden 18
dygn.
Fig. 5 Ruvfläcken hos hane D, fotograferad 27 april 1952, då ungarna var 4 dagar gamla.
The incubation patch of the male D, when the young were 4 days old.
Antalet ägg beroende ar födotillgången.
Till nyssnämnda siffror bör göras den kommentaren, att proportionen mellan 3- och 4-äggkullar ovan är påverkad av att studienötkråkorna utfodrades med nötter under söndagar vintertid. De fyra 3-äggkullarna tillhörde par, som åtminstone de åren ej subventionerats med nötter eller annan föda. Antalet ägg i nötkråkans kull är alltså klart beroende av tillgången till föda under häckningstiden, d.v.s. normalt av hur mycket nötförråd, som nötkråkorna kunnat upplägga föregående höst.
Hur initieras effektiv ruvning hos hanen?
Nötkråkan, liksom våra andra fåglar, erhåller fysiologiskt genom hormon förmågan att ge äggen rätt ruvningstemperatur i och med utbildning av ruvfläcken (Jones 1971 m.fl.). Att honan applicerar rätt värme just från och med sista ägget, kan ges en enkel fysiologisk - etologisk förklaring. Men en fascinerande, obesvarad fråga är: hur vet hanen, att han skall börja aktiv ruvning just. vid tredje ägget i en 3-kull men först med fjärde ägget i en 4-kull? Det ligger nära till hands att antaga, att honan på något sätt signalerar starten genom sitt beteende.
Anmärkning.
Uppgifterna ovan har till största delen och med mera utförligt siffermaterial publicerats tidigare. Då detta emellertid skett i en festskrift, som i ornitologiska kretsar torde ha begränsad spridning (se Swanberg 1956), då denna saknar svensk sammanfattning och undersökningen skett inom Skövde Fågelklubbs geografiska region, har redaktionen för Grus föreslagit den nu gjorda sammanfattningen. Se ovan
Litteratur
Bailey, R.E. 1952. The incubation patch of passerine birds. Condor, 54: 121-136.
Becker, P. & Ringleben, H. 1975. Welche Bewandtnis hat es mit dem "Brutfleck" beim hane des Zitronenfinken? J.F.O. 116: 325-326.
Erpino, M.J. 1969. Seasonal cycle of reproductive physiology in the Black-billed magpie. Condor 71: 207-279.
Holstein V 1942 Duehogen. Köpenhamn 1950. Spurvehogen. Köpenhamn.
Huggins, R.A. 1941. Egg temperatures of wild birds under natural conditions. Ecology 22: 148-157.
Jones, R. E. 1965. The incubation patch of birds. Biol Rev 46: 315-339.
Kolthoff, K. 1934. Några ord om fåglarnas ruvfläckar. Fauna o Flora 29:49-55.
Lange, B. 1928. Die Brutffecke der Vögel und die für sie wichtigen Hauteigentümlichkeiten. Gegenbaurs Morph Jahrb. 59:601-712.
Lloyd, J.A. 1965. Seasonal development of the incubation patch in the starling (Sturnus vulgaris). Condor 67:67--72.
Mewaldt, L.R. 1952. The incubation patch of the Clark nutcracker. Condor 54:361.
Mewaldt L R 1956. Nesting behavior of the Clark nutcracker. Condor 58:3-23.
Nice, M. M. 1954. Problems of incubation periods. Condor 56: 173-197.
Ryves, B.H. 1929. Variability in incubation- and fledgingperiods. Brit Birds 22: 203-205. Stresemann, E. 1934. Aves. I Kükenthal, W: Handbuch der Zoologie, Bd 7, Teil 2. Berlin.
Swanberg, P O 1950. On the concept of incubation period. Vår Fågelv. 9:63-80.
Swanberg, P O 1956. Incubation in the Thick-billed nutcracker, Nucifraga c. caryocatactes. I Bertil Hanström - zool.papers in honour of his sixty-fifth birthday, s 278-297.
Swanberg, P O 1976. Nötkråkans Nucifraga c. caryocatactes, vana att upplägga vinterförråd. Grus 2: 10-18.
Tucker, B. W. 1943. Brood patches and the physiology of incubation. Brit Birds 37:22-28.
Incubatlon in the Thick - billed Nutcracker, Nucifraga c. caryocatactes.
Summary: The paper deals with research findings, previously published 1956. The Nutcracker broods the eggs continuously and steadily from laying of the first egg, protecting them but without giving them true incubation temperature. The laying interval between the eggs of a clutch is slightly more than 24 hours. Incubation starts with the last egg laid and, as a rule, all eggs hatch in one day. Incubation period is 18 days. The male shares incubation with the female, in the forenoon from sunrise untill 12.00 it takes 46 - 48% of the incubation (9 nests). In observations at two nests from sunrise until sunset the males incubated on an average 34% of the time (26 and 43% respectively), cf. fig. 2. The female incubates in the night. The male has a true incubation patch of the same size and structure as in the female. This develops rapidly in the nest - building time. As the male changes from brooding to true incubation with the third egg in 3-egg clutches and with the fourth egg in 4 - eggs clutches, it is suggested that the female by its behaviour incluces the male to change the brooding behaviour to incubation.
Territory in thick billed Nutcracker.
By P O Swanberg Received on 9 January 1956. [From Ibis, 98 (1956): 412-419.]

The general habits of the Thick-billed Nutcracker have been studied by me in Sweden since 1940. For this study an area was chosen in the conifer forest of Billingen, a hill in Central Sweden. In this area up to 5 pairs were continuously studied between 1941 and 1953. The nesting was also checked in 1954 and in 1955. Of the original birds one male remained in May 1955, which then proved to be at least 15 years old, and there is good evidence of the breeding of the same bird in the same territory in 1940 and in 1939, when the nest was found. Thus it is thought to be 18 years old or more.
The plateau of the hill is covered with spruce forest, and extensive coppices consisting mostly of hazel - bushes are to be found on the slopes and on the surrounding ground. The above mentioned area is part of the spruce forest of the central and northern sector of the hill, and the total of the conifer forest of this sector is estimated to be about 38 square km. (9000 acres). In a census carried out in 1944 - 46, the area of each Nutcracker pair was mapped as completely as possible. In all 145 pairs were counted, well dispersed over the forest, and the results of the census were checked by samples taken at random in 1947.
I have studied various aspects of the habits of the Nutcracker in many other parts of Sweden, and certain details of the food - storing habit, too, in some islands of the Åland archipelago (in the Baltic between Sweden and the mainland of Finland) as well as in Pinus cembra habitat in the Engadine Switzerland, in May and June 1954.
The Nutcracker is a food specialist. Always nesting in conifer wood, the Scandinavian population nevertheless subsists on a good supply of hazel (Corylus avellana) within reach of the nesting site. In the autumn nuts are carried from the hazel - coppices and stored in the conifer wood up to 6 km. (4 miles) away (Swanberg 1951).
In other areas, where Pinus cembra occurs, e.g. in the Engadine, the Nutcracker both collects and stores its winter food in the nesting habitat. Some birds, indeed, are seen carrying cembra nuts in their throat - pouch far away as much as 10 - 12 km. from the collecting place (Sutter & Amann 1953).
Slender billed Nutcrackers also feed during the whole winter on their stores of cedar nuts (Formosof 1933).
Thick - billed Nutcrackers are truly resident birds and my observations indicate that they are paired for life. Each pair has its special area in the conifer wood and, so far as my experience goes, they hold their area, when they have found a suitable one, for life. In three cases males have, after losing their females, retained their territories and introduced new mates into them.
The territory boundaries are difficult to determine. Four territories that could be drawn with satisfactory exactitude measured 11.2 - 14.9 hectares or, on an average, 13.2 hectares (32.7 acres). The greatest density I know is on Sottunga, an island of the Åland archipelago, where 8 - 11 September 1949 Nutcrackers equivalent to 40 pairs were observed laying up stores, each pair in its particular area or territory. These stores were all fairly well spread over a forest area of about 360 hectares. That is in reality less than 9 hectares (22.2 acres) for each pair. The relation between the number of adult birds and that of immature birds is, indeed, not known. The unusually great density may be explained in two ways - the surrounding water might have a partly restraining influence upon the emigration of young birds and another and interesting factor might be the absence of groundliving rodents in the woods of the island, with, as a consequence, less need of protection for the numerous food stores.
To the area that I call "territory", the members of the Nutcracker pair carry nuts throughout the autumn, as long as nuts are available. The winter stores are used not only outside the breeding season but also for raising the young. In their territory the mates spend the whole of the winter, each pair in its own territory, as long as the ground is covered with snow. The birds are able to find their stores even under the deepest snow, and neither snow nor severe cold drive them out of a territory which is once established. Within the boundaries of the territory all the nestsite selection goes on and, consequently, the nest is always built in the territory where the stores were laid up.
When the ground of the slopes is bare of snow and the breeding season approaches, Nutcrackers make excursions of a kilometer or more, evidently completing the food. The principal food of the young in the nest consists of nut kernels from the winter stores. Supplementary food for the young is brought partly from inside the territory but to a considerable degree from outside it.
After leaving the nest the young stay for two or three weeks in the territory, or at least with the territory as a base for their short excursions in no man's land. Throughout the summer months, until the beginning of the ripening of the nuts, the young accompany their parents every morning to the deciduous woods far away from the home in the conifer forest, but towards evening the family returns to roost in its own territory. Nutcrackers help their young to get food for a very long time. I have seen them feeding young 105 days old in the territory; 9 days later young were begging and, judging from the characteristic sounds, were then fed by one of the parents.
When the time for food storing has come, family bonds are broken by the young, which disperse. Six Swedish Nutcrackers, ringed as nestlings, have been recovered after periods of ½ - 5½ years at 7.5 - 40 km. (5-25 miles) from their birthplace.
 Thumbnail. the bird in nest.
Thumbnail. the bird in nest.
Advertising the territory
In spite of the fact that each pair of Nutcrackers keeps its own area these areas do not correspond satisfactorily to the generally accepted definition of territory as an area "defended against individuals of the same species" if by "fighting" we mean real fighting. Such fighting is rarely seen or never, nor is a consistent and clear defence of the territory against intruders by means of sounds or posturing found. Nevertheless, each pair obviously has its definite territory. As an example my Nutcracker pair A was proved to have kept the same territory for at least fourteen years. When the old female was lost the male retained its territory for the next breeding season with its new mate. In all I have found at least 19 nests of A in the same territory, but although A has always been surrounded by other pairs, other Nutcrackers, as far as I know, have never made an attempt to nest in the same area.
Partly because Nutcrackers pair for life and the territories are kept for a very long time, the formation of the sexual bond and the establishment of a new territory are very difficult to observe in the Nutcracker. There is therefore a possibility that the birds defend their territory with actual fighting during the short time of pairing - up and establishing a new territory. Once well advertised by the owners and well known by the neighbours, a territory kept for the bird's lifetime obviously does not need so much defending as a short lived territory.
Territory is not proclaimed by the song, which is evidently only of sexual character (Swanberg 1951), but probably by the bird's harsh, farreaching cries. After a period of strict silence in winter and as a sign of the beginning of the breeding cycle, the birds start calling simultaneously in the mornings a short time before nestbuilding. They preferably call along the boundaries facing the favourite places of the neighbouring Nutcrackers, and call most just before nestbuilding. When the eggs are laid absolute silence sets in again, and during the incubation time the forests, to many bird observers, appear to have been emptied of Nutcrackers.
In the summer months much calling is heard again from every territory, but the significance of the calls at this period when the young are with the parents in the territories, is difficult to state.
During the next period, when they begin to gather nuts and when newcomers probably have to look for and occupy areas suitable for foodstoring as well as for breeding, Nutcrackers call comparatively frequently and, if the territory owners are at home, other Nutcrackers passing in flight over the territory are often saluted with calls, even if they show no sign of alighting.
I suggest that the essential point is that, with or without encounters, the other Nutcrackers respect the ownership of the territoryholders or, in the words of Lack (1954), "that in both territorial and colonial species, dispersion is primarily due to the avoidance of occupied or crowded ground by potential settlers, not to the aggressive behaviour of those in occupation".
Behaviour towards intruders
As already stated, other Nutcrackers do not try to settle in established territories but they can visit or trespass on such areas without being chased out. An interesting fact is that on many occasions ceremonial gatherings of several Nutcrackers (often 8 - 10) take place within certain territories immediately before and during nestbuilding with the territoryowners as friendly watchers or even as participants. It is difficult to give a satisfactory explanation of these puzzling events, but I would suggest that a primary, practical function of these gatherings is to facilitate the meeting of unpaired birds that are not going to breed until the next spring. If so, the party might consist partly of well established territoryholders (which offer no threat to others), partly of young birds that are not going to settle until four or five months later on.
When nuts are placed experimentally within a territory both the owners chase away intruders, often violently, if they attempt to share that special food. This usually occurs without calling and the attacks, even when violent, do not aim at driving the intruders outside the boundaries of the territory.
Other Details
As to a difference in territorial behaviour between mated and unmated birds, it is very difficult to give general conclusions. However, in several instances unmated birds in the breeding season have been recorded as being without territorial behaviour. On one occasion the sex was determined - the bird was a female that replaced the lost previous mate of my male A the next year.
The most interesting observation on non breeding birds was made in connection with an emigration of Thickbilled Nutcrackers that took place in 1949. Some of these arrived on the island of Öland, east of southern Sweden. They found a comparatively good supply of nuts and stayed, in spite of the fact that this island with its abundance of hazel has at best only very poor conifer woods of the type required by the species for breeding. In a small wood of planted pines stores were laid up in September and, when I visited the place on 16 October, I noted that 10 - 25 Nutcrackers had their haunts in the wood. After watching from before the awakening of the birds I noted: "There seemed to be no territory claiming cries in the morning". Judging from the sounds, all the birds were young ones. Next year I spent the days 17 - 19 March, 30 April and 1 May there and noted no signs of nesting. Some Nutcrackers, however, were apparently paired, though they did not breed. Two months later, 24 June, 12 Nutcrackers were counted in the wood, and at the same time I noted: "It seems to me that the Nutcrackers beginning the second year of their life now, after the normal nesting season and after emptying the nutstores, do not hold territories, but are rather flocking to a common roosting place à la Hawfinch at Billingen and others". Between 24 June and 11 September they all disappeared. That wood was apparently not suitable for nesting. I concluded that the Nutcrackers made off to seek a good nesting ground and to establish territory elsewhere, if possible.
In my experience no other species here in Scandinavia has such similar requirements that it could be regarded as a serious competitor in the territory. The Jay (Garrulus glandarius) is attacked in the vicinity of the nest but not in all parts of the territory and not outside the nesting cycle of the Nutcracker.
The Goshawk (Accipiter gentilis), where it lives, is no doubt for the Nutcracker the most dangerous predatory bird. I have seen Nutcrackernestlings in the hawk's nest. Some unexplained nestrobbing is thought to be the work of the Marten (Martes Martes). As protection from these two predators the spacing out obviously can be of some positive value.
Does territorial behaviour set an upper limit to the number of pairs present in a definite area, or does it merely space out the pairs present whose number in the area as a whole has been primarily determined by other factors ? There is good evidence that in districts with a limited supply of hazel but with abundance of breeding ground the number of territories is limited by the amount of hazel. On the other hand, if things are inverted, as on Kinnekulle, a hill 27 - 28 km. from Billingen where a small conifer wood is surrounded by hazel in abundance, the number of territories is evidently reduced. Only about 10 Nutcracker territories are to be found at Kinnekulle in spite of the extensive supply of hazel (N.G. Karvik, personal communication). I think it justifiable to regard territorial behaviour as the instrument that has served to give the small conifer wood of Kinnekulle only a normal density of Nutcrackers and to prevent overcrowding.
The Nature and value of Nutcracker territory
If advertising by calling can be regarded as "fighting", I think the territory of the Nutcracker corresponds to the usual definition of territory during certain periods of the year. In other periods, however, the birds are silent, inactive, not to say retiring. Is the territory then, really, to be regarded as a "territory" all the year ? In my opinion it is.
The function of the territory, as I see it, may certainly be to secure an area where other pairs do not interfere with nesting or with storing food for a subsequent breeding season; "fighting" and advertising might be considered merely as means for securing this function in the periods within which it can be seriously threatened. In my opinion the territory of the Nutcracker is a "breeding territory". It is true that every pair spends the winter in its own territory, but the breeding is apparently the primary factor, and the situation and the size of the territory, together with the arranging of the stores, may have for its goal the successful rearing of offspring.
The most conspicuous aspect of the territory of the Nutcracker in Scandinavia is as a storage area for food, carried by the birds from rather distant collecting grounds. Yet the primary factor may be the tendency of the bird to secure a territory in a habitat that is suited for nesting (as well as for stores), and the bird's tendency to store the special food within the selected area may be secondary. When this special food, as in Scandinavia, is available only far away from the territory, carrying flights have to be made over a long distance. The above mentioned 145 pairs of central and northern Billingen collected nuts in common feeding areas, but they stored the food in 145 different areas of the vast conifer forest. In other words: the distribution of the stores to the territories is evidently of vital importance for the bird.
As advantages of storing the special food in the territory the following can be suggested:
- (a) The stores are concentrated in a limited area, known by the owner in every detail. This evidently facilitates selecting numerous and good storeplaces as well as remembering these places the next year.
- (b) The habit guarantees accessibility of the stores in the breeding time, a most important factor, as it gives larger clutchsize, on the average. If the amount of the special food available for the young is good, the size of the clutch is normally 4 eggs and more young are raised. When the special food is lacking, the clutch is always reduced to 3 eggs. Thus, birds storing the food in the territories must have come to predominate over others. Furthermore, the stores are easily accessible, which means that the bird has the advantage of shorter flights, with saving of time and a better opportunity to keep watch over its family.
- (c) Interference with other pairs is avoided. If the stores had not been distributed to all the well scattered territories, a probable effect and danger could have been a tendency to restrict the storing to more limited parts of the forest, to a limited zone at a distance more representative for the species than, as now, for the individual bird. In all probability such a habit would have to a greater extent exposed the stores to various dangers from competitors of the same species, from rodents and so on. I have seen a Nutcracker watch another bird's storing and, when left alone, empty the store. Experiments have shown that voles or (and) mice are able to find nuts hidden by Nutcrackers in the ground.
Consequently the following qualities of the territory might be of survival value:
- (1) The territory makes rational food storing possible, i.e. it preserves the special food for the birds in winter as well as in the breeding season. (There is no clear evidence that a certain amount of supplementary - not stored - food within the boundaries of the territory is of decisive importance.)
- (2) The territory secures to the Nutcracker pair a place of refuge with which they are familiar, thus giving a maximum of protection against predators, cold weather, etc.
- (3) The territory gives shelter from predators also by spacing the pairs out.
- (4) The territory helps to maintain the bond between the mates. This is, indeed, a highly hypothetical suggestion.
To sum up in one sentence: I would suggest that the territory of the Nutcracker has as its function to give the mates a common refuge, well known in every detail and thus, during the breeding period as well as in winter, as perfect and protective as possible; and to prevent, by spacing out, a possible reduction or removal of these positive qualities of the territory by overcrowding. The general lines of this suggestion agree, in my opinion, with the views of Tinbergen (1952) on a territory of such different form as that of the Herring Gull (Larus argentatus) and with the opinions of Lack (1954) on gregarious nesting, solitary nesting and dispersion. Yet the Nutcracker territory has an extraordinary connection with food, too, as it provides shelter not only to the birds themselves but also to their most important food.
Summary
1. The main features of the territory in the Thickbilled Nutcracker are:
- (a) the territory is kept for life and inhabited throughout the year;
- (b) the special food, nuts, is stored in the territory;
- (c) song, copulation, nest site selection and nestbuilding take place only in the territory;
- (d) territories examined were 11.2 - 14.9 hectares, averaging 32.7 acres, believed to be the normal size in Sweden.
2. There is no clear evidence that the territory has the function of securing more than a certain amount of food other than stored.
3. The territory is respected by other Nutcrackers as to its main functions, while simple visits by other Nutcrackers and ceremonial gatherings in the territory are tolerated.
4. Both territory holders attack intruders that try to take nuts but as they do not chase them out the behaviour is not regarded as truly territorial behaviour.
5. The territory is proclaimed by the calls, the song being exclusively sexual.
6. Unmated birds have not been recorded with territorial behaviour.
7. When the special food is limited and the nesting habitat ample, the population is limited by the amount of the special food; but when the special food is abundant and the nesting habitat limited, overcrowding seems to be prevented by territorial behaviour.
8. It is suggested that the function of territory in the Nutcracker is to give the holders a wellknown refuge from predators, to serve as a food storing place, and by spacing out to prevent these positive qualities from reduction by overcrowding.
References.
FORMOSOF, A. N. 1933. The crop of cedar nuts, invasions into Europe of the
Siberian Nutcracker (Nucifraga caryocatactes macrorhynchos Brelun) and
fluctuations in numbers of the Squirrel (Sciurus vu 1-aris L.) J. Anim. Ecol. 2:
70-81.
LACK, D. 1954. The natural regulation of animal numbers. Oxford.
SUTTER, E. & AMANN, F. 1953. Wie weit fliegen vorratssammelnde
Tarmenhäher? Orn. Beob. 50: 89-90.
SWANBERG, P. 0. 1951. Food storage, territory and song in the Thickbilled
Nutcracker. Proc. 10 Int. Orn. Cong. 1950 : 545-554.
TINBERGEN, N. 1952. On the significance of territory in the Herring Gull.
Ibis 94: 158-159.
Food Storage, Territory and Song in the Thick - billed Nutcracker
Per Olof Swanberg
The thick - billed nutcracker (Nucifraga caryocatactes caryocatactes) is found in southern and central Sweden up to the river Dalälven, 80 km north of Uppsala. A careful investigation of all known breeding localities in Sweden and Finland in the course of a study of the bird's habits has shown that the population of the nutcracker in Scandinavia, where Pinus cembra does not occur, must subsist on the supply of hazel-nuts (Corylus avellana) in the vicinity of the nesting site.
Hazel-nuts, however, are normally obtainable in nature only in a few autumn months. The nuts fall to the ground during October, and in the absence of nutcrackers they are mostly eaten or carried away by small rodents before winter. But as early as the end of August the nutcrackers begin to store up nuts, which they carry in their throat-pouch from the hazel-coppice to some place more or less deep in a coniferous wood. Their habit of carrying and storing the nuts of hazel or Pinus cembra has been known for a long time in different parts of Europe, and their life during two or three autumn months is actually completely devoted to food storage.
When the work begins in earnest, the nutcrackers leave their haunts in the spruce forest early in the morning, generally before sunrise, and fly direct to the hazel-coppice, which is often several and may he as much as six kilometers away. Within an hour of sunrise many have returned, loaded with nuts. These are stored in small heaps of different sizes, sometimes only three or four, often eight or nine, in the ground. The stores are laid at a depth of a bill's length, or under the lichen or moss cover of a rock. The first picture, which is from one of the islands of the Åland archipelago, shows such a store. I watched the nutcracker through my glasses and saw how and where it hid its nuts, and later I went to the spot. Although I knew exactly where the nuts lay, it was impossible to find anything on the surface which betrayed their position. But by turning the lichen cover I exposed them to view (Fig. 1). The heap consisted of eighteen nuts, which had been neatly arranged in two layers like sardines in a tin.

Fig. 1. A nutcracker - store consisting of 18 nuts. The lichen cover is turned over. Only the upper layer is visible. (The picture is from Åland in Finland).
When the birds are engaged in storing, they do not stay longer in the forest than is necessary for hiding the nuts. Three minutes after they arrive, on an average, they return to the hazel-coppice. The work proceeds fairly regularly throughout the day. By way of an example I have marked in a diagram (Fig. 2) the number of times nuts have been carried by several nutcrackers which I watched during one of many observation days.
Fig. 2. Nucifraga c. caryocatactes. Billingen, Central Sweden. Number of flights of birds carrying nuts in throat-pouch per hour, observed from a fixed point in territory A, September 26, 1948. Between sunrice and sunset.
As you see, the birds were very busy all the day. They began to arrive with nuts seventeen minutes after sunrise and stopped only at sunset.
The nutcrackers carry nuts throughout the autumn as long as nuts are available. The frequency of their journeys was studied in different years. Diagram № 2 (Fig. 3) is from 1948 when the crop of hazel-nuts was a rich one.
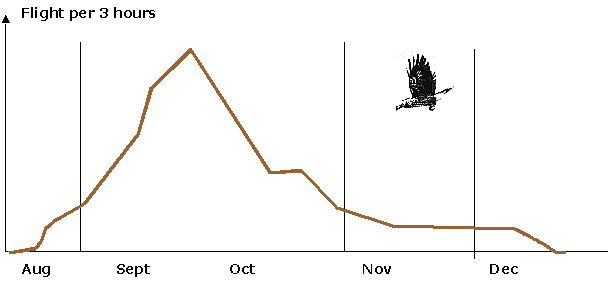
Fig. 3. Nucifraga c. caryocatactes. Billingen, Central Sweden. Frequency of food storing. Number of flights, with nuts in throat-pouch, observed in three-hours periods between sunrise and noon, Aug.-Dec., 1948, from a fixed point in territory A.
It is based on observations from a fixed point and calculated from the number of journeys observed in three-hour periods. The top of the curve shows that storing activity is at a maximum when the nuts are ready to drop. It will be seen that during a great part of the year, normally about three months, the nutcracker works for its future subsistence from morning to evening with an intensity which can be compared with that of man.
Such a pronounced habit of intensive and specialized work could hardly have developed if the nutcracker had not succeeded in making use of its stores. Carl von Linné (1748-52: 94), in his lectures here in Uppsala two hundred years ago, said that the nutcracker "as the farmer says takes the clouds as a mark" when it hides its nuts, and so will not be able to find them again.
Many more recent writers have made similar assumptions. Yet ornithologists have recorded some instances of a nutcracker finding its stores again (Studer & Fatio, 1901: 314, and others).
In point of fact in the spring the nutcracker still has such a large supply of hazel-nuts that the principal part of the food of the young in the nest consists of nut kernels. For it has a fantastic power of rediscovering its stores. Every day in winter it digs out nuts from one or more hiding places. The stores may be snow-covered, sometimes for several months, but this does not deter the birds, for they can find their stores even under eighteen inches of snow.
One photograph shows a typical example of a store that has been dug out by a nutcracker (Fig 4).

Fig. 4. Excavation in the snow where the nutcracker has dug out its nuts.
Heinroth's (1924-28: 259) experiments with the raven (Corvus corax) and my own with the nutcracker have shown that the bird cannot conceivably have been aided by the sense of smell. On winter days which offered optimum conditions of observation I have counted the number of excavations in the snow with nutshells in or near them, and also the number of excavations where there were no signs of the bird having found its nuts. Out of 351 such excavations which I have examined, the nutcracker had succeeded in finding its nuts in 86 per cent. This figure is in fact considered to be too low as an index of the retentive power of the nutcracker's memory.
It is of special interest that almost one-third of these records occurred in each of the months, January, February and March (in the March group I have included the few April records), and that the percentage of successful excavations in snow is practically the same for all three months, viz. 86 per cent. This indicate that the nutcracker has a kind of abstract recollection, which, figuratively speaking, helps it in remembering which stores were used earlier in the winter. Thus it practically never searches for a store already emptied, even if new snow has fallen.

Fig 5 The small depression in the ground out of which six nuts were dug under the snow cover shows that the nutcracker did not have to seek for the nuts.
Another picture (Fig. 5) shows the small depression in the ground out of which six nuts were dug. It is evident from the appearance of the excavation that the bird did not have to seek for the nuts.
Each pair has its special area in the coniferous wood. In my experience, after ten years' study, nutcrackers hold their area, when they have found a suitable one, for life.
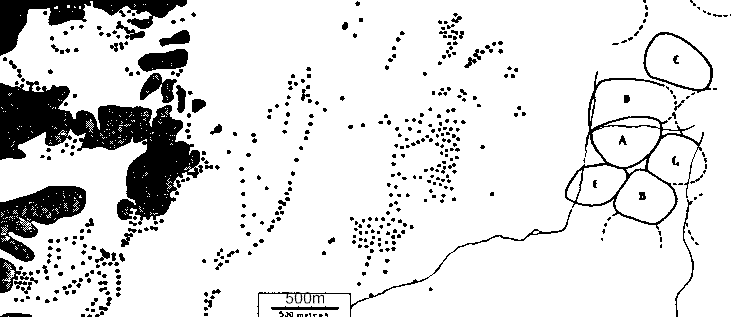
Fig. 6. Living area of some pairs of nutcrackers. Billingen, Central Sweden. To the right the territories in the spruce forest. In the middle and to the left is communal feeding area. Each black spot marks a group of hazels.
The sketch-map (Fig. 6) shows the living area beyond the boundaries of which my best-known pairs of nutcrackers will never, or at any rate only for a short distance, move. To the right is the spruce forest on Billingen mountain, then comes the slope of the mountain covered by deciduous wood. To the left is the low-lying, partly cultivated country. Each black spot marks a group of hazels. On the right I have drawn the approximate boundaries of the areas on the plateau-top of the mountain which are occupied by the different pairs of nutcrackers. These areas average 13 - 14 hectares. The rest is communal feeding area.
In my notes I have always called these special areas `territories'. But in more careful descriptions I have often used more cautious terms, on account of the well-known definitions of territory given by Mayr (1935), Noble (1939), N. Tinbergen (1936, 1939), Nice (1943) and others, which claim that a territory should be defended "by a fighting bird against individuals of the same species and sex" or the like.
In spite of intensive studies of nutcrackers for many years, I have never observed any true territorial fights or any consistent defence of the territory against intruders by means of sounds or posturing. Nevertheless, each pair obviously has its defined territory. As an example, my nutcracker pair A are proved to have held the same territory for at least ten years. During this time I have found fourteen nests of A in it, but although A has always been surrounded by other pairs, other nutcrackers have never made any attempt to nest in the same area. Broadly speaking, the areas are used only by their holders, but it is not unusual for other nutcrackers to trespass and rest in them without being attacked, and sometimes they may even pay friendly visits to each other, as during the ceremonial gatherings, which take place early in the morning immediately before or during nest-building. During these gatherings neighboring birds meet pair A in A's area, probably without any practical purpose and without the owners making any attempt to chase them out of their territory.
It should, however, be added that nutcrackers pair for life, and that their territory corresponds in its general type to that of such birds as the chaf finch (Fringilla coelebs), the reed-bunting (Emberiza schoeniclus), the snow-bunting (Plectrophenax nivalis), which Howard called "breeding territory" and N Tinbergen "sexual territory". These sexual territories, according to Tinbergen and others, are defended "shortly before and during the formation of a sexual bond", and for several reasons the formation of the sexual bond in the nutcracker is very difficult to observe. Thus I cannot deny the possibility that the nutcracker may in fact defend its territory with actual fighting during the short time of pairing up and establishing a new area. On the other hand Mary Erickson (1938) has pointed out that in the wren-tit (Chamaea fasciata), which holds territory continuously throughout the year and retains the same territory for a number of years, probably for life, "fighting was observed in all months of the year". Moreover, Angus Robertson (1949: 301) has pointed out that in south-western Australia many birds defend a territory to a certain extent all the year round, and that, for instance, butcherbirds (Cractias) and kookaburras (Dacelo gigas), which "defend permanent territories in pairs ... will foregather at times even during the breeding season, with neighboring members of the species, . . . well within one or the other pair's territory without friction".
It is, however, a fact that every pair of nutcrackers disposes of a special area throughout the year, and that the right of possession of this area is respected by other nutcrackers without actual territorial fights. Thus this area can hardly be described by the ornithological technical term 'territory', although more than most bird-territories it corresponds to what human beings mean by territory, and in spite of the fact that its purpose corresponds to that indicated by Howard (1920) when he wrote that "securing a territory is (then) part of a process which has for its goal the successful rearing of offspring".
In defining territory some authors consider it wrong to include any reference to the function of territory, as the function may be, and in fact is, so different for different species. Thus the territorial fights or the mere symptom of establishing the territory are regarded as the prime consideration, and such definitions of territory
It should, however, be added that nutcrackers pair for life, and that their territory corresponds in its general type to that of such birds as the chaf finch (Fringilla coelebs), the reed-bunting (Emberiza schoeniclus), the snow-bunting (Plectrophenax nivalis), which Howard called "breeding territory" and N Tinbergen "sexual territory". These sexual territories, according to Tinbergen and others, are defended "shortly before and during the formation of a sexual bond", and for several reasons the formation of the sexual bond in the nutcracker is very difficult to observe. Thus I cannot deny the possibility that the nutcracker may in fact defend its territory with actual fighting during the short time of pairing up and establishing a new area. On the other hand Mary Erickson (1938) has pointed out that in the wren-tit (Chamaea fasciata), which holds territory continuously throughout the year and retains the same territory for a number of years, probably for life, "fighting was observed in all months of the year". Moreover, Angus Robertson (1949: 301) has pointed out that in south-western Australia many birds defend a territory to a certain extent all the year round, and that, for instance, butcherbirds (Cractias) and kookaburras (Dacelo gigas), which "defend permanent territories in pairs ... will foregather at times even during the breeding season, with neighboring members of the species, . . . well within one or the other pair's territory without friction".
It is, however, a fact that every pair of nutcrackers disposes of a special area throughout the year, and that the right of possession of this area is respected by other nutcrackers without actual territorial fights. Thus this area can hardly be described by the ornithological technical term 'territory', although more than most bird-territories it corresponds to what human beings mean by territory, and in spite of the fact that its purpose corresponds to that indicated by Howard (1920) when he wrote that "securing a territory is (then) part of a process which has for its goal the successful rearing of offspring".
In defining territory some authors consider it wrong to include any reference to the function of territory, as the function may be, and in fact is, so different for different species. Thus the territorial fights or the mere symptom of establishing the territory are regarded as the prime consideration, and such definitions of territory approximate rather to definitions of the use of the technical term 'territory'. But this leaves us with no general designation for the phenomenon of such vital importance to the bird, which the breeding territory in a deeper sense represents and which is expressed by each pair securing a territory, although we may call this phenomenon "breeding territory," "sexual territory" or the like when fights occur as indications of this function.
In my opinion it is, however, justifiable also to regard as breeding territory an area like that of the nutcracker, which is the property of a single pair of birds in a considerably stricter sense than, for instance, that of a chaffinch (Fringilla coelebs). Although it is not defended in the true sense of the word, and intruders are not chased away, the territory is proclaimed throughout a great part of the year. It applies to the nutcracker as well as to the wren-tit (Chamaea fasciata) that "the general impression is not of birds continuously exerting pressure or sparring for an opening, but rather of their acceptance of the existing status" (Erickson 1938: 263).
The purpose of territory in the nutcracker seems to be to secure every pair a certain area of their own, which they get to learn in every detail and which thus ensures particularly good shelter for the adult birds, their nest and offspring, and which above all is suitable for well scattered and adequate stores for the winter, which are of vital importance at any rate for the populations of Nucifraga caryocatactes caryocatactes which, in Scandinavia, have to feed on hazel-nuts.
The function of proclaiming the position and boundaries of the territory, which is ascribed to the song of many territorial birds, is probably exercised by the harsh cries of the nutcracker, audible for a long distance. These cries have many functions, but one of them seems to be the proclamation of the territory. This is indicated partly by the behavior of the birds, partly by the frequency of the cries.
The song, on the contrary, is weak and can hardly serve as advertising song. As song I do not consider the many different notes which are mainly heard during ceremonial gatherings before nesting and in late summer. In my opinion the actual song consists of the various piping, whistling and whining notes, which are uttered rhythmically, more or less regularly interspersed with distinct clicking sounds, and by the male only. During the song the bird takes up a position with its bill upwards and the throat dilated. Singing from the top of a tree, however, is rather abnormal, and the bird usually sings in a concealed position at a low height above the ground. Even this supports the view that it is not an advertising song indicating the territory for rivals or attracting a mate.
l have only heard the song during a few weeks before and during nest building. Often the male interrupts nest-building for a short sung. Of the different functions which have been ascribed to bird-song in general (Armstrong 1942, 1947: 294), the only one which seems to apply to the nutcracker is that of facilitating the synchronization of the male and female sexual cycles. Armstrong says that another function of song is to indicate the stage the bird has reached in its sexual cycle. This of course can also apply to the song as an independent function, but in my opinion in the nutcracker it only forms part of the above-mentioned facilitating of the synchronization. In other words, it is a mere mating song, which despite its pronounced subdued character must not he confused with sub-song. It is interesting to compare conditions in Australia, where according to Robinson (1947: 11; 1949: 293) many birds are resident territory-holders, and mating songs are therefore prevalent and of vital importance for them.
Nötkråkans Parningsdans
P O har i sitt intensiva umgänge med kråkan upptäckt att de kan ha en parningsdans, som han dokumenterat spåren av. Ingen har emellertid någonsin sett detta. I flera år har jag försökt få P O att berätta mer därom samt visa bilden, men P O hade sådana krav på vad han gav ut. Vi pratade mycket om det, och här har jag funnit två bilder, som jag inte vill undanhålla dig.


These thubnail may be the first documents about the Nutcracers
playground. Nobody has ever seen it nor did Swanberg.
PO har sedan 1938 studerat Nötkråkan här på Billingen med sina närmare 200 par. Han bjöd in Nisse Lindman och tidningar att skriva om detta.
1954-05-15 kommer GP på besök och berättar att det finns planer på en bok. Tyvärr kom aldrig någon bok, då Hornborgasjön kom att uppta all hans tid och kraft. Med nu tillgång till facit där Arn och turismkommersen tagit sjön i anspråk kan det finnas anledning att fundera kring om det var värt 50 års uppoffring på nötkråkans bekostnad.
1955-04-26 har han inbjudit en tidning som får följa med till nötkråkans skog. Här berättas att hanen måste vara 18 år och att han tagit en ny hona sedan första hustrun tydligen dukat under under föregående år. Här berättas att kråkan på sitt 14 ha stora revir kan ha upp till 1000 förråd och att släktingen smalnäbbade nötkråkan vid felslagen Cembratall fruktgivning tvingas att flytta. De kråkor som Swanberg hade var i det närmaste tama och att han kunde locka på de.
Ett meddelande i Fauna och Flora 1938
PO skriver att han återfunnit en liten population 1938 där Gustaf Kolthoff fann dem 40 - 50 år tidigare. Redan 1935 och 1936 fann han häckande par och 1938 tar han följande välkända bild från ett bo 1 maj 1938.

Detta var nog startskottet för hans studier av denna art som han blev så nära bekant med.
Smalnäbbad Nötkråka
Av och till invaderar den sibiriska kusinen, vilket skedde hösten 54 med den påföljden att ett på Gotska Sandön kvardröjande par blev föremål för fotografering.

Smalnäbbad under häckningsperioden 1955.